You might also like
- Signature Whistles Italy Terranova Et Al. 2021Document12 pagesSignature Whistles Italy Terranova Et Al. 2021jenn.aereaNo ratings yet
- Geographic Distribution and Taxonomy of the Chipmunks of WyomingFrom EverandGeographic Distribution and Taxonomy of the Chipmunks of WyomingNo ratings yet
- History and Biology of The Black ProteusDocument34 pagesHistory and Biology of The Black ProteusGregor AljancicNo ratings yet
- Mazzoil Et AlDocument10 pagesMazzoil Et Alapi-3828346No ratings yet
- tmpFA4F TMPDocument8 pagestmpFA4F TMPFrontiersNo ratings yet
- Abundance and Densities of Beaked and Bottlenose Whales (Family Ziphiidae)Document8 pagesAbundance and Densities of Beaked and Bottlenose Whales (Family Ziphiidae)Cécile PersohnNo ratings yet
- Population Status Update For The Northern Resident Killer Whale IN 2018Document13 pagesPopulation Status Update For The Northern Resident Killer Whale IN 2018王道远No ratings yet
- Sex Ratios of Loggerhead Sea Turtles Caretta Caretta During The Juvenile Pelagic StageDocument13 pagesSex Ratios of Loggerhead Sea Turtles Caretta Caretta During The Juvenile Pelagic StagealejoolduNo ratings yet
- Reptilian All The Way?: LetterDocument1 pageReptilian All The Way?: LetterJamesComeyJustaBitchNo ratings yet
- RS469 SI09 Cephalorhynchus 1988Document354 pagesRS469 SI09 Cephalorhynchus 1988battwomanNo ratings yet
- Residency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanDocument11 pagesResidency and Abundance of Sperm Whales (Physeter Macrocephalus) in Nemuro Strait, Hokkaido, JapanGianni PavanNo ratings yet
- Beebe Cooper 2002Document12 pagesBeebe Cooper 2002etik ainun rohmahNo ratings yet
- FSR 175Document10 pagesFSR 175Michaiel PiticarNo ratings yet
- Repertorio Sonoro de Los DelfinesDocument20 pagesRepertorio Sonoro de Los DelfinesFélix Armadá JuárezNo ratings yet
- Blue Zone The AKEA StudyDocument8 pagesBlue Zone The AKEA StudyMiddioPNo ratings yet
- Observations of Localised Movements and Residence Times of Wobbegong Sharks (Orectolobus Halei) at Fish Rock, NSW, AustraliaDocument10 pagesObservations of Localised Movements and Residence Times of Wobbegong Sharks (Orectolobus Halei) at Fish Rock, NSW, AustraliaismailsalaydrusNo ratings yet
- Nautilus: Biology, Systematics, and Paleobiology As Viewed From 2015Document17 pagesNautilus: Biology, Systematics, and Paleobiology As Viewed From 2015Casa Manu Vale SEMOLANo ratings yet
- de Pinna, M. A New Family of Neotropical Freshwater Fishes From Deep Fossorial Amazonian Habitat, With A Reappraisal of Morphological Characiform Phylogeny (Teleostei Ostariophysi)Document31 pagesde Pinna, M. A New Family of Neotropical Freshwater Fishes From Deep Fossorial Amazonian Habitat, With A Reappraisal of Morphological Characiform Phylogeny (Teleostei Ostariophysi)Iván Sipión GuerraNo ratings yet
- First Report On The Density and Size of Napoleon WrasseDocument13 pagesFirst Report On The Density and Size of Napoleon WrasseGoresan RimbaNo ratings yet
- Preproposal FinalDocument9 pagesPreproposal Finalapi-254424729No ratings yet
- Capture-Recapture Estimates of Hector'S Dolphin Abundance at Banks Peninsula, N E W ZealandDocument13 pagesCapture-Recapture Estimates of Hector'S Dolphin Abundance at Banks Peninsula, N E W ZealandFathimah Dini HanifahNo ratings yet
- Off Shores DalheimDocument11 pagesOff Shores DalheimBudi AfriyansyahNo ratings yet
- Marine Biology Research: A B A BDocument14 pagesMarine Biology Research: A B A BKaritoNo ratings yet
- Hybridism Rissos Dolphins Bottlenose DolphinsDocument7 pagesHybridism Rissos Dolphins Bottlenose DolphinsXyz AnonimNo ratings yet
- Filinia Species in Brackish Waters at St. Maarten (Dutch Caribbean)Document6 pagesFilinia Species in Brackish Waters at St. Maarten (Dutch Caribbean)Hidrobiologia 1 IA SASNo ratings yet
- Carpophilus Keys For Identification of Carpophilus PDFDocument41 pagesCarpophilus Keys For Identification of Carpophilus PDFErica Livea100% (3)
- Abyssal Fauna of Polymetallic Nodule Exploration Areas, Eastern Clarion-Clipperton Zone, Central Pacific Ocean: Amphinomidae and Euphrosinidae (Annelida, Amphinomida)Document42 pagesAbyssal Fauna of Polymetallic Nodule Exploration Areas, Eastern Clarion-Clipperton Zone, Central Pacific Ocean: Amphinomidae and Euphrosinidae (Annelida, Amphinomida)Andrey RozenbergNo ratings yet
- 203 FullDocument27 pages203 FullProf YoupaNo ratings yet
- Case Study: Hippocampus Spp. Project Seahorse: 1. 1.1 Scientific and Common NamesDocument19 pagesCase Study: Hippocampus Spp. Project Seahorse: 1. 1.1 Scientific and Common NamesEkhaPutriNagFisheryNo ratings yet
- Word Materi 9Document13 pagesWord Materi 9santiNo ratings yet
- Wenzel Et Al. 2020 Humpback Whales in The Cape Verde Islands Migratory Patteerns Resightings and Abundance Aquatic MammalsDocument11 pagesWenzel Et Al. 2020 Humpback Whales in The Cape Verde Islands Migratory Patteerns Resightings and Abundance Aquatic MammalsMicéal FlynnNo ratings yet
- Today: Chris Gleed - Owen, CGO EcologyDocument13 pagesToday: Chris Gleed - Owen, CGO Ecology3582No ratings yet
- Enc 1101 - Project 1 Final Version - Gabriela MorenoDocument7 pagesEnc 1101 - Project 1 Final Version - Gabriela Morenoapi-742553212No ratings yet
- Butterflies of SulawesiDocument265 pagesButterflies of Sulawesikim7382No ratings yet
- Mating Behavior and Rapid Concealing of Nuptial Coloration in Males of The Fishgod Blenny Malacoctenus Ebisui (Teleostei: Blenniiformes)Document5 pagesMating Behavior and Rapid Concealing of Nuptial Coloration in Males of The Fishgod Blenny Malacoctenus Ebisui (Teleostei: Blenniiformes)CRISTIAN MOISES GALVAN VILLANo ratings yet
- SiphonophoresDocument12 pagesSiphonophoresNicolasNo ratings yet
- Oray 2015Document3 pagesOray 2015Hector RodriguezNo ratings yet
- Two New Damsel y Species From Polillo Island, Philippines (Odonata: Platystictidae)Document6 pagesTwo New Damsel y Species From Polillo Island, Philippines (Odonata: Platystictidae)Geelord Dela CruzNo ratings yet
- IndotestudoDocument5 pagesIndotestudoNina RosdianaNo ratings yet
- 058 2016 Ferrando Et Al Polar BiologyDocument13 pages058 2016 Ferrando Et Al Polar BiologyharisankarhsNo ratings yet
- Janik Et Al 2014 - Cetacean Vocal Learning and CommunicationDocument6 pagesJanik Et Al 2014 - Cetacean Vocal Learning and CommunicationVini ReveilleauNo ratings yet
- Morphospeccies: Prepared By: Chris Olemar A. LlidoDocument17 pagesMorphospeccies: Prepared By: Chris Olemar A. LlidoCE YMNo ratings yet
- Animal BiotelemetryDocument10 pagesAnimal BiotelemetryLindsay DurkinNo ratings yet
- The Cardiidae Mollusca Bivalvia of TropiDocument90 pagesThe Cardiidae Mollusca Bivalvia of TropimuricidaeNo ratings yet
- Book - Gruper of The WorldDocument11 pagesBook - Gruper of The WorldLidiya KirilovaNo ratings yet
- Reproductive Parameters of The Amazon River Dolphin or Boto, Inia Geoffrensis (Cetacea: Iniidae) An Evolutionary Outlier Bucks No TrendsDocument11 pagesReproductive Parameters of The Amazon River Dolphin or Boto, Inia Geoffrensis (Cetacea: Iniidae) An Evolutionary Outlier Bucks No TrendsNataliaaa684No ratings yet
- Fin Whale Acoustic Presence and Song Characteristics in Seas To The Southwest of PortugalDocument16 pagesFin Whale Acoustic Presence and Song Characteristics in Seas To The Southwest of PortugalGianni PavanNo ratings yet
- CAT 2020 Question Paper With Solutions Slot 1 Bodhee PrepDocument68 pagesCAT 2020 Question Paper With Solutions Slot 1 Bodhee Prepmanaswini sharma B.G.100% (1)
- Defran and Weller 1999Document15 pagesDefran and Weller 1999api-3828346No ratings yet
- The Dinoflagellate Dinophysis Norvegica Biological and Ecological Observations in The Baltic SeaDocument10 pagesThe Dinoflagellate Dinophysis Norvegica Biological and Ecological Observations in The Baltic SeaHabib Ben HabibNo ratings yet
- Marine VisionDocument16 pagesMarine VisionYuliana PurnamasariNo ratings yet
- CAT 2020 Question Paper With Answer Keys Slot 1 Bodhee PrepDocument33 pagesCAT 2020 Question Paper With Answer Keys Slot 1 Bodhee PrepAROKYASWAMY LNo ratings yet
- Pac Sci Early View 71 2 7Document15 pagesPac Sci Early View 71 2 7Marine BiologyNo ratings yet
- WHB Polarbear Pop 2011Document8 pagesWHB Polarbear Pop 2011sarah0rogersNo ratings yet
- Rankin 2005Document6 pagesRankin 2005api-3828346No ratings yet
- Artikel Filum MolluscaDocument10 pagesArtikel Filum MolluscaListiyani Putri AzzahroNo ratings yet
- Global Trade in Illicit Elephant IvoryDocument59 pagesGlobal Trade in Illicit Elephant IvoryBeatriz TurNo ratings yet
- Living Planet 2014Document180 pagesLiving Planet 2014Beatriz TurNo ratings yet
- Forests and Land UseDocument1 pageForests and Land UseBeatriz TurNo ratings yet
- Indians in BrazilDocument50 pagesIndians in BrazilBeatriz Tur100% (1)
- The NobodiesDocument1 pageThe NobodiesBeatriz TurNo ratings yet
- You Should Think Twice Before Buying Tuna If You Love SharksDocument1 pageYou Should Think Twice Before Buying Tuna If You Love SharksBeatriz TurNo ratings yet
- Dirty Fuels Clean Futures Report 2014Document32 pagesDirty Fuels Clean Futures Report 2014Beatriz TurNo ratings yet
- Poisonous PoliciesDocument12 pagesPoisonous PoliciesBeatriz TurNo ratings yet
- The Spirit LevelDocument345 pagesThe Spirit LevelBeatriz TurNo ratings yet
- Create Your Dream LifeDocument113 pagesCreate Your Dream LifeBeatriz Tur100% (3)
- 11 Arguments Against CaptivityDocument3 pages11 Arguments Against CaptivityBeatriz TurNo ratings yet
- 24 01 11 Happiness Challenge FinalDocument12 pages24 01 11 Happiness Challenge FinalinspgadgetNo ratings yet
- Child Poverty InsightsDocument3 pagesChild Poverty InsightsBeatriz TurNo ratings yet
- Vegetarian Starter Guide (MFA)Document17 pagesVegetarian Starter Guide (MFA)Vegan Future50% (2)
- 365 Daily QuotesDocument32 pages365 Daily QuotesAna SilivestruNo ratings yet
- Wapnick Kenneth A Course in MiraclesDocument1,754 pagesWapnick Kenneth A Course in MiraclesCecilia PetrescuNo ratings yet
- Palm Oil ListDocument7 pagesPalm Oil ListBeatriz TurNo ratings yet
- 15 Ways To Boost Your ConfidenceDocument19 pages15 Ways To Boost Your ConfidenceBeatriz Tur100% (1)
- Guidelines On How To Behave When Diving With SharksDocument9 pagesGuidelines On How To Behave When Diving With SharksBeatriz TurNo ratings yet
- Esclavage Moderne Rapport de L'organisation Walk FreeDocument136 pagesEsclavage Moderne Rapport de L'organisation Walk FreeMo GattereauNo ratings yet
- Report On Further Comparisons of Morgan Acoustic RepertoireDocument9 pagesReport On Further Comparisons of Morgan Acoustic RepertoireBeatriz TurNo ratings yet
- 64 Daily Practices ShortDocument11 pages64 Daily Practices ShortBeatriz Tur100% (1)
- Companies That Do Test On AnimalsDocument15 pagesCompanies That Do Test On AnimalsBeatriz TurNo ratings yet
- David Kirby's Marie Claire Article, "Enemy Waters," Regarding A SeaWorld Trainer's DeathDocument3 pagesDavid Kirby's Marie Claire Article, "Enemy Waters," Regarding A SeaWorld Trainer's DeathJeffrey Ventre MD DCNo ratings yet
- 10 Essential Facts About RhinosDocument13 pages10 Essential Facts About RhinosBeatriz TurNo ratings yet
- Javan Rhino SpeciesDocument1 pageJavan Rhino SpeciesBeatriz TurNo ratings yet
- Sumatran Rhino SpeciesDocument1 pageSumatran Rhino SpeciesBeatriz TurNo ratings yet
- White Rhino SpeciesDocument1 pageWhite Rhino SpeciesBeatriz TurNo ratings yet
- Black Rhino SpeciesDocument1 pageBlack Rhino SpeciesBeatriz TurNo ratings yet
- Grade5 Unit1Document5 pagesGrade5 Unit1api-239494748No ratings yet
- Dog Training Methods ReviewDocument48 pagesDog Training Methods Reviewnagorelasfuentes100% (1)
- Harry PotterDocument9 pagesHarry PotterRiaan RaoNo ratings yet
- The 12 Days of ChristmasDocument2 pagesThe 12 Days of ChristmasCarla Muss-JacobsNo ratings yet
- Psychology of ThinkingDocument19 pagesPsychology of ThinkingDiyana HalimNo ratings yet
- NCP Impaired Gas ExchangeDocument2 pagesNCP Impaired Gas ExchangeGabriel Tolentino70% (10)
- Artificial Salmon SpawningDocument28 pagesArtificial Salmon SpawningjimborenoNo ratings yet
- List of SlaughterHouses in UAEDocument2 pagesList of SlaughterHouses in UAEDr-Muhammad Awais Ch100% (1)
- Psychophysiological Detection of Deception Analysis Ii - Course #503Document67 pagesPsychophysiological Detection of Deception Analysis Ii - Course #503John PrasetioNo ratings yet
- Carnage Island A Rejected Mate Standalone Romance Reject Island by Lexi C PDFDocument347 pagesCarnage Island A Rejected Mate Standalone Romance Reject Island by Lexi C PDFZowiwi wi100% (4)
- ?weebee Beginners Bunny CALDocument17 pages?weebee Beginners Bunny CALyovanna ortizNo ratings yet
- Reading Comprehension For Grade 3 PDFDocument6 pagesReading Comprehension For Grade 3 PDFIVAN CONRADNo ratings yet
- Principles of Selection and Mating SystemsDocument29 pagesPrinciples of Selection and Mating SystemsUnknown StudentNo ratings yet
- Product Catalogue 2010Document112 pagesProduct Catalogue 2010Dennis NguyenNo ratings yet
- Behavior of Feeding in Guppy Poecilia ReticulataDocument6 pagesBehavior of Feeding in Guppy Poecilia ReticulataresearchanimalsciencesNo ratings yet
- Daily Reading Book 1 PDFDocument11 pagesDaily Reading Book 1 PDFSanjeevan SivapaleswararajahNo ratings yet
- Toto The Turtle Wid OtherDocument43 pagesToto The Turtle Wid OtherleetolbeanNo ratings yet
- Comparative Evaluation Between F1 Duroc HampshireDocument9 pagesComparative Evaluation Between F1 Duroc HampshireGed L'EsparverNo ratings yet
- WALIKI Mens Knits 2010Document17 pagesWALIKI Mens Knits 2010Waliki Design TeamNo ratings yet
- Anatomy (Chapter - More MCQS) Solved MCQs (Set-3)Document4 pagesAnatomy (Chapter - More MCQS) Solved MCQs (Set-3)Saleem AliNo ratings yet
- Harcourt GK Phonics Practice Book StudentDocument144 pagesHarcourt GK Phonics Practice Book StudentІрина КрупаNo ratings yet
- Neonatal Sepsis: Ekawaty Lutfia Haksari Perinatology, Department of Child Health Gadjah Mada UniversityDocument28 pagesNeonatal Sepsis: Ekawaty Lutfia Haksari Perinatology, Department of Child Health Gadjah Mada Universityireneaurelia100% (1)
- Hymns of OrpheusDocument71 pagesHymns of OrpheusJustice DubhghaillNo ratings yet
- Creatures & Constructs A Monster Manual Supplement For EberronDocument74 pagesCreatures & Constructs A Monster Manual Supplement For EberronJason BryantNo ratings yet
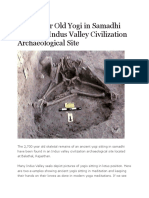
- 2,700 Year Old Yogi in Samadhi Found in Indus ValleyDocument12 pages2,700 Year Old Yogi in Samadhi Found in Indus ValleyMisterNo ratings yet
- Association Between Caesarean Delivery and Isolated Doses of Formula Feeding in Cow Milk AllergyDocument6 pagesAssociation Between Caesarean Delivery and Isolated Doses of Formula Feeding in Cow Milk Allergyirene aureliaNo ratings yet
- DR Mushtaq Amna Ashraf Aqsa Bashir Ayesha Bibi Hira Nisar Iqra HabibDocument5 pagesDR Mushtaq Amna Ashraf Aqsa Bashir Ayesha Bibi Hira Nisar Iqra HabibtaibiNo ratings yet
- Wuthering HeightsDocument284 pagesWuthering Heightslamija7No ratings yet
- NCGG Irish Festival Grand Marshals 2017Document1 pageNCGG Irish Festival Grand Marshals 2017NewzjunkyNo ratings yet
- North Sea GeologyDocument170 pagesNorth Sea Geologyoturens100% (1)
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessFrom EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo ratings yet
- Merle's Door: Lessons from a Freethinking DogFrom EverandMerle's Door: Lessons from a Freethinking DogRating: 4 out of 5 stars4/5 (326)
- The Other End of the Leash: Why We Do What We Do Around DogsFrom EverandThe Other End of the Leash: Why We Do What We Do Around DogsRating: 5 out of 5 stars5/5 (65)
- Will's Red Coat: The Story of One Old Dog Who Chose to Live AgainFrom EverandWill's Red Coat: The Story of One Old Dog Who Chose to Live AgainRating: 4.5 out of 5 stars4.5/5 (18)
- The Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationFrom EverandThe Dog Listener: Learn How to Communicate with Your Dog for Willing CooperationRating: 4 out of 5 stars4/5 (37)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueFrom EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueRating: 5 out of 5 stars5/5 (3)
- Roxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueFrom EverandRoxane Gay & Everand Originals Presents: Good Girl: Notes on Dog RescueRating: 4.5 out of 5 stars4.5/5 (24)
- Come Back, Como: Winning the Heart of a Reluctant DogFrom EverandCome Back, Como: Winning the Heart of a Reluctant DogRating: 3.5 out of 5 stars3.5/5 (10)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- An Eagle Named Freedom: My True Story of a Remarkable FriendshipFrom EverandAn Eagle Named Freedom: My True Story of a Remarkable FriendshipNo ratings yet
- Show Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredFrom EverandShow Dog: The Charmed Life and Trying Times of a Near-Perfect PurebredRating: 3.5 out of 5 stars3.5/5 (13)
- Dogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowFrom EverandDogland: Passion, Glory, and Lots of Slobber at the Westminster Dog ShowNo ratings yet
- The Wrong Dog: An Unlikely Tale of Unconditional LoveFrom EverandThe Wrong Dog: An Unlikely Tale of Unconditional LoveRating: 4.5 out of 5 stars4.5/5 (26)
- Tamed and Untamed: Close Encounters of the Animal KindFrom EverandTamed and Untamed: Close Encounters of the Animal KindRating: 3.5 out of 5 stars3.5/5 (32)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- Puppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!From EverandPuppy Training 101: How to Train a Puppy, Training Your Own Psychiatric Service Dog, A Step-By-Step Program so your Pup Will Understand You!Rating: 5 out of 5 stars5/5 (85)
- Arthur: The Dog Who Crossed the Jungle to Find a HomeFrom EverandArthur: The Dog Who Crossed the Jungle to Find a HomeRating: 4.5 out of 5 stars4.5/5 (20)
- Edward's Menagerie: Dogs: 50 canine crochet patternsFrom EverandEdward's Menagerie: Dogs: 50 canine crochet patternsRating: 3 out of 5 stars3/5 (5)
- Your Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesFrom EverandYour Dog Is Your Mirror: The Emotional Capacity of Our Dogs and OurselvesRating: 4 out of 5 stars4/5 (31)
- Arthur: The Dog who Crossed the Jungle to Find a HomeFrom EverandArthur: The Dog who Crossed the Jungle to Find a HomeRating: 4.5 out of 5 stars4.5/5 (16)
- MINE!: A PRACTICAL GUIDE TO RESOURCE GUARDING IN DOGSFrom EverandMINE!: A PRACTICAL GUIDE TO RESOURCE GUARDING IN DOGSRating: 4 out of 5 stars4/5 (15)
- The Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenFrom EverandThe Faraway Horses: The Adventures and Wisdom of America's Most Renowned HorsemenRating: 4.5 out of 5 stars4.5/5 (49)
- How To Speak Dog: Mastering the Art of Dog-Human CommunicationFrom EverandHow To Speak Dog: Mastering the Art of Dog-Human CommunicationRating: 4 out of 5 stars4/5 (66)