Professional Documents
Culture Documents
Morfologia y Estructura Bacteriana
Uploaded by
stinkipsCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Morfologia y Estructura Bacteriana
Uploaded by
stinkipsCopyright:
Available Formats
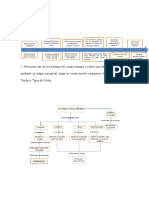
MORFOLOGIA Y ESTRUCTURA BACTERIANA Las bacterias o procariotas, son microorganismos unicelulares que se reproducen por fisin binaria (divisin
simple). Muchos tienen vida libre. Contienen informacin gentica, sistemas de produccin de energa y sistemas biocinticas necesarias para el crecimiento y reproduccin. La gran divisin de los seres vivos se realiza segn el tipo de clulas: eucariota y procariota, tambin poseen este tipo de clulas algas, hongos y protozoos. Las bacterias son microorganismos unicelulares, siendo entonces clulas procariotas. Existen dos grupos de procariotas evolutivamente distintos, las eubacterias y las arquebacterias. Una clula bacteriana tpica tiene las siguientes estructuras: Material gentico ADN, bajo forma de un cromosoma nico que no est rodeado de membrana nuclear, esta caracterstica es la diferencia fundamental con la clula eucariota, la cual posee siempre membrana nuclear. Presentan adems ribosomas, citoplasma y membrana. SERES VIVOS: Organizacin unicelular sin diferenciacin Clula procariota (ncleo primitivo) Procariota: bacterias Organizacin unicelular o colonial Clula eucariota (ncleo verdadero) Protistas: protozoarios, fitoflagelados Multicelulares. Diferenciacin celular en tejidos Clula eucariota Reinos: animal y vegetal Un o multicelular Un o multinuclear Clula eucariota Hongos, algas citoplasmtica. Por fuera est la pared bacteriana, estructura que por su composicin bioqumica se puede decir que es propia de las bacterias, ya que las clulas vegetales tienen una pared celular pero est compuesta por celulosa. Algunas bacterias como los micoplasmas, carecen de pared celular. Al decir estructuras variables queremos decir que estas estructuras existen en algunas bacterias y no en todas, y aun en un mismo grupo bacteriano o una misma cepa bacteriana las puede presentar o no, dependiendo de las condiciones en donde se desarrolle. Las estructuras variables no son necesarias para la vida de la clula bacteriana. Tamao: Las bacterias presentan una amplia diversidad de tamaos, que va desde 0.5 a 2 micrmetros y algunas pueden llegar a 10 micras. No son visibles por supuesto al ojo humano y se visualizan con microscopio ptico (MO) o electrnico (ME). Las bacterias pueden ser observadas al MO sin ser coloreadas si se las coloca en glicerol o soluciones no acuosas que aumenten el ndice de refraccin. Las bacterias se pueden observar sin colorear utilizando la tcnica de microscopa de campo oscuro en la que utilizando un condensador especial se ven sobre un fondo oscuro como cuerpos brillantes. Esta tcnica se usa para el examen de microorganismos no teidos en suspensiones lquidas como es el caso de Treponema pallidum, agente de la sfilis, que se observan como espiroquetas delgadsimas. Forma: Al MO o ME las bacterias se presentan con una morfologa definida que est determinada por su pared rgida. Se pueden presentar como esfricas, ovaladas, denominndose cocos. Si la forma es cilndrica se denominan bacilos o
bastones. Estos bastones pueden ser rectos, curvos o con forma de espiral, en este ltimo caso les llamamos espirilos. Las clulas bacterianas pueden mantenerse unidas en grupos despus de que se han dividido, pero conservando siempre la independencia una clula de otra. Cocos o bacilos pueden agruparse en cadenas, en el caso de los cocos, cuando se presentan as agrupados, se denominan estreptococos. Tambin se pueden presentar como diplococos. Si los planos de divisin son variados pueden agruparse en ttradas o como racimos, denominndose estafilococos. Los bacilos pueden ser muy cortos, recibiendo el nombre de cocobacilos, otras veces pueden ser muy largos, pudiendo tener una longitud 10 veces superior a su dimetro. Los extremos pueden ser redondeados o rectos, pueden presentarse aislados, en largas cadenas o pueden agruparse en empalizadas o formando letras chinas. La morfologa bacteriana se observa al MO sin tincin como se seal, o utilizando diferentes tcnicas de tincin que ayudan a su mejor visualizacin ya que son incoloras. Las diferentes tcnicas de tincin consisten en colorear las clulas con diferentes colorantes que tienen afinidad por materiales celulares especficos. Hay colorantes catinicos, de carga positiva que tienen afinidad por constituyentes celulares de carga negativa, como los cidos nucleicos y los polisacridos. Algunos ejemplos de colorantes catinicos: azul de metileno, cristal violeta, safranina. Ms adelante analizaremos algunos detalles de la coloracin ms utilizada en Bacteriologa, denominada coloracin de Gram en honor a quien en 1884 la describi. Es esta una coloracin diferencial que permite dividir a las bacterias en Gram positivas, refirindose a aquellas que toman el primer colorante utilizado que tie las bacterias de color violeta y las Gram negativas, las que toman el ltimo colorante utilizado en la tcnica como colorante de contraste que las tie de rojo o rosado. Este tipo de coloracin se denomina diferencial por lo que acabamos de analizar. Otras coloraciones simples como el azul de metileno, se utilizan para observar simplemente la morfologa. Analizaremos la clula desde su interior para luego dedicarnos a las estructuras externas y apndices. Las estructuras internas de la clula estn inmersas en el citoplasma, solucin acuosa y viscosa, que contiene solutos orgnicos e inorgnicos y elementos especializados como ribosomas y grnulos de inclusin. Material gentico: Ribosomas: La clula bacteriana presenta ribosomas libres en el citoplasma con coeficiente de sedimentacin de 70S a diferencia de la clula eucariota que es de 80S. Se presentan tambin como polirribosomas que son cadenas de ribosomas asociados a ARN mensajero y en parte en relacin con el ADN cromosmico. Contienen todos los componentes que permiten la sntesis proteica. Las clulas poseen ms ribosomas si estn creciendo en medios ricos. El alto contenido de ARN determina gran afinidad por los colorantes bsicos.
ADN bacteriano: Como se seal, la clula procariota a diferencia de la eucariota carece de una membrana nuclear, tampoco posee nuclolo, ni aparato mittico, y nunca configura una masa cromosmica definida. El material gentico est compuesto de una estructura fibrilar, constituida por un ADN circular de doble cadena, enrollado sobre s mismo. Si bien se asocia a protenas bsicas, estas no son verdaderas histonas. Los mesosomas son, al parecer, invaginaciones de la membrana citoplasmtica y participan en la divisin celular y en la replicacin de ADN. Si bien hay autores que opinan que son artefactos de tcnica que se observan en las microfotografas electrnicas, hay otros que han observado que claramente el ADN se fija al mesosoma en la etapa previa de la divisin celular. En las ltimas dcadas se han desarrollado muchas tcnicas que permiten estudiar el ADN bacteriano, todas ellas con importantes aportes en la investigacin y el diagnstico. Por ejemplo, es posible por tcnicas de secuenciacin, conocer genomas enteros de bacterias o sectores de l que tengan inters, ya sea porque codifican la produccin de alguna estructura, una toxina, etc. Estos sectores de ADN, se pueden separar del ADN total con enzimas de restriccin para su posterior anlisis o utilizacin en estudios de biologa molecular. Estas tcnicas de biologa molecular estn destinadas a identificar sectores de ADN especfico de una bacteria, por ejemplo: las tcnicas de hibridacin que identifican un trozo de ADN con una sonda gentica complementaria de ese sector y marcada con un compuesto revelador; o la tcnica de reaccin en cadena de la polimerasa (PCR) que amplifica miles de veces el sector de inters. Plsmidos y episomas: Algunas bacterias poseen material gentico extra cromosmico, denominados plsmidos y episomas. Los plsmidos son elementos genticos constituidos por secuencias de ADN cortas circulares que se replican en forma autnoma. Como se ver en el captulo de gentica bacteriana, stos poseen genes que codifican factores de agresin, factores de resistencia a antibiticos, produccin de toxinas, etc. Los episomas son elementos genticos extra cromosmicos que pueden existir en forma autnoma o incorporada al material gentico. Membrana celular: Estructura delgada que rodea a la clula de 8 nm de espesor. Es una estructura vital, si se altera, la clula pierde su vitalidad. Composicin: es similar a la mayor parte de las membranas biolgicas. Una doble capa fosfolipdica en donde el cido graso hidrofbico se orienta hacia el interior y el glicerol hidroflico hacia el exterior de la clula. A diferencia de la clula eucariota no posee esteroles (excepto los micoplasmas). Algunos antibiticos, cuyo blanco de ataque en la clula bacteriana es la membrana celular, actan inmediatamente que la clula se pone en contacto con stos. Los antibiticos de este tipo, son por su mecanismo de accin, txicos para los tejidos humanos y poco utilizados en medicina clnica. Funcin: Delimita el interior del exterior celular, es una barrera osmtica muy importante, interponindose a la libre difusin entre el medio ambiente interno y externo. Es una membrana con permeabilidad selectiva, a travs de la cual ingresan los nutrientes y salen los productos de desecho. El transporte de solutos se realiza
muchas veces por un sistema de transporte activo, como se analizar al estudiar el metabolismo bacteriano. En la membrana celular estn los sistemas de fosforilacin, oxidacin y transporte de electrones (citocromos) para la produccin de energa. Dentro de las diversas actividades enzimticas vinculadas a las protenas de la membrana citoplasmtica, podemos sealar su participacin en la biosntesis del ADN, de los constituyentes de la pared, etc. Ya hemos hecho mencin ms arriba de los mesosomas que son organelos que se asocian a la membrana. Se observan al ME como sacos citoplasmticos que contienen estructuras verticiladas, laminares, tubulares o vesiculares, y a menudo se asocian con tabiques de septacin. Se ha demostrado que los mesosomas fijan el ADN cromosmico a la membrana citoplasmtica. Pared celular: Estructura rgida presente como ya se dijo en la mayora de las bacterias, se sitan por fuera de la membrana citoplasmtica. Es una estructura vital para las bacterias que las poseen. Si sta se destruye o se impide su formacin, la clula pierde su viabilidad. El espesor de la pared en los diferentes microorganismos, oscila entre 0,150 m y 0,500 m de (m=micra). Podemos imaginar entonces la pared celular, como una macromolcula gigante constituida por peptidoglicano, que en forma de bolsa rodea toda la clula, representando del 10 al 40 % del peso de la bacteria. Unido al N-acetil murmico se encuentra un tetrapptido como se aprecia en la figura 4. Los tetrapptidos de una cadena de peptidoglicano se unen con los de la otra a travs de puentes peptdicos. Los aminocidos que forman los tetrapptidos son los siguientes: L-Alanina, Acido DGlutmico, Acido mesodiaminopimlico (o LLisina) y D5 Alanina. Podramos resumir de la siguiente manera la sntesis del pptidoglicano: En el momento de la divisin celular se debe formar la nueva pared celular. Las autolisinas, enzimas producidas por la propia bacteria, forman brechas en la "vieja pared" de la clula en divisin. A nivel de esas brechas o aberturas es donde se agrega el peptidoglicano de la nueva pared en formacin. La mayor parte de la informacin se ha obtenido de la sntesis del peptidoglicano de Staphylococcus aureus y la explicacin que sigue se restringir a este organismo. En el citoplasma se une el N-acetil Murmico a un transportador lipdico de membrana: el bactoprenol, luego se une la Nacetil Glucosamina y, finalmente, el puente de pentaglicina. El bactoprenol transporta el bloque formado a travs de la membrana citoplasmtica. Una vez en el espacio periplsmico, estos bloques son colocados en las brechas ya formadas. El paso final y fundamental para una correcta funcin de la pared es la unin los monmeros de peptidoglicano entre si. Esta unin la realizan enzimas denominadas transpeptidasas. A este paso se lo conoce como transpeptidacin. La Penicilina y sus derivados, los antibiticos ms usados en Medicina, actan precisamente inhibiendo la sntesis de la pared celular, llevando a la lisis de la bacteria. La unin de los puentes peptdicos se pueden hacer como ocurre en la mayora de las bacterias por uniones directas transpetdicas entre la D-Alanina de un puente peptdico y el cido diaminopimlico (DAP) de otro tetrapptido adyacente. En Staphylococcus aureus la reaccin de transpeptidacin se efecta mediante un
puente de pentaglicina, el cual une la L-Lysina de un tetrapptido con la D-Alanina de otro tetrapptido adyacente. Existen diferencias en el espesor de esta estructura bsica, el peptidoglicano de las bacterias Gram positivas tienen una capa gruesa de 0,02 a 0,06 m en forma de multicapas, mientras que las bacterias Gram negativas y las bacterias cido alcohol resistentes la tienen ms fina, de 0,01 m. Al peptidoglicano se le unen otros componentes que son, por lo tanto, tambin integrantes de la pared. Son diferentes en las bacterias Gram positivas, negativas y cido alcohol resistentes. Revisaremos algunos aspectos de la composicin de los tres tipos de paredes. Pared de bacterias Gram negativas: Si observamos la pared de las bacterias Gram negativas al microscopio electrnico podemos observar 3 zonas: 1. La membrana citoplasmtica. 2. El espacio periplasmtico, ubicado entre la membrana citoplasmtica y el peptidoglicano. 3. Una fina capa de peptidoglicano. 4. Membrana externa que contiene fosfolpidos, el lipopolisacrido (LPS) caracterstico de estas bacterias y protenas (protenas de membrana externa). Esta capa est estrechamente unida al peptidoglicano y se la considera un componente de la pared celular de las bacterias Gram negativas. En el lipopolisacrido se pueden distinguir 3 componentes diferentes bioqumicamente. La porcin lipdica, el lpido A, est inmersa en el centro de la membrana externa. En su sector externo la porcin polisacardica, el denominado polisacrido O, o antgeno O, est ubicado en la cara externa. Entre estos dos componentes est el core del LPS, que es tambin polisacardico. El antgeno O es muy variable en su composicin entre las diferentes familias y especies de bacterias Gram negativas, en cambio, el polisacrido del core es constante en las diferentes bacterias Gram negativas. El polisacrido O por su variabilidad es utilizado frecuentemente para la clasificacin serolgica de familias bacterianas de inters mdico. Al lipipopolisacrido (LPS) se le denomina endotoxina, siendo el lpido A su la porcin txica. Como el lpido A est inmerso en el centro de la membrana slo ejerce sus efectos txicos cuando la clula es lisada, ya sea por ataque del complemento y fagocitosis, o por lisis celular como consecuencia de la accin de antibiticos. Si se inyecta a un animal de experimentacin endotoxina, ocurren una serie de eventos fisiolgicos, el primero es la fiebre. La endotoxina estimula a las clulas del husped a liberar protenas denominadas pirgenos y stos modifican el centro termorregulador del encfalo. Pero el animal tambin puede sufrir otras alteraciones como disminucin de leucocitos y plaquetas en sangre, sufriendo finalmente un estado inflamatorio generalizado que lo puede llevar a la muerte (fenmeno de Shwartzman-Sanarelli). Efectos similares ocurren cuando por destruccin bacteriana hay endotoxinas en sangre. La gravedad del cuadro clnico se sabe hoy que depende en parte de la cantidad de endotoxinas circulantes, pudiendo determinar desde un simple cuadro infeccioso con fiebre, a sepsis, shock sptico o la muerte.
La toxicidad del lpido A radica primariamente en su habilidad para activar el complemento y estimular la liberacin de citoquinas por parte de las clulas del husped (leucocitos, macrfagos, etc.). La membrana externa contiene adems numerosas protenas. Algunas de ellas la atraviesan de lado a lado, constituyendo canales, denominados porinas. Estos poros permiten el pasaje de molculas de bajo peso molecular, como nutrientes. Las grandes molculas no atraviesan la membrana externa, por esto estas bacterias, a diferencia de las Gram positivas, no son sensibles a una enzima, la lisozima que destruye el peptidoglicano. Incluso algunos antibiticos de alto peso molecular no pueden atravesar la membrana externa y, por lo tanto, las bacterias no son sensibles a ellos. Pared de las bacterias Gram positivas: Lo primero a sealar es la gruesa capa de peptidoglicano en forma de mltiples capas, los cidos lipoteicoicos estn embebidos en la membrana citoplasmtica por su porcin lipdica. Los cidos teicoicos tienen por funcin estabilizar la pared. La superficie externa del peptidoglicano de las bacterias Gram positivas est usualmente cubierta de protenas. Los diferentes grupos de bacterias Gram positivas y las diferentes especies, difieren en la composicin de sus protenas y cidos teicoicos, siendo esto til para la clasificacin serolgica y la identificacin. En la pared de las bacterias Gram positivas no existe una endotoxina, sin embargo, la presencia de estas bacterias en los tejidos y en la sangre determina sntomas similares al shock sptico que ocurre ante la presencia de endotoxinas. Las mismas citoquinas que se liberan ante la presencia de los LPS, se sabe que son liberadas ante presencia de la pared de las bacterias Gram positivas con sus mismos efectos. A la luz de las investigaciones actuales, al aparecer fragmentos de peptidoglicano, cido teicoicos o una combinacin de los dos, juegan un papel semejante al LPS. Si se inyectan a animales tienen efectos similares que los LPS. Al peptidoglicano de la pared de las bacterias Gram positivas, se pueden unir protenas como la protena M de Streptococcus pyogenes (o Streptococcus grupo A) que se vincula con la virulencia, y tambin se pueden asociar polisacridos como el polisacrido C que permite la clasificacin en serotipos del gnero Estreptococos.
You might also like
- BARTONELOSISDocument32 pagesBARTONELOSISestudperNo ratings yet
- Apuntes MicrobiologiaDocument45 pagesApuntes MicrobiologiaEduardo MorelosNo ratings yet
- Ci - ECOLOGIA SEM 1 A 8Document68 pagesCi - ECOLOGIA SEM 1 A 8RonaldoQuiñonezPaucarNo ratings yet
- Practica 1 Columna de Winogradsky 170402192055Document11 pagesPractica 1 Columna de Winogradsky 170402192055maria pintoNo ratings yet
- Línea de Tiempo de Aportes A La Teoría CelularDocument4 pagesLínea de Tiempo de Aportes A La Teoría Celularluisa fernanda mesa barbosaNo ratings yet
- Preguntas-EBAU-Introducción A La CélulaDocument5 pagesPreguntas-EBAU-Introducción A La CélulaInés García GarcíaNo ratings yet
- Exp BiorremediacionDocument26 pagesExp BiorremediacionLilian Andrea100% (1)
- CUESTIONARIO 3 MicrobiologiaDocument6 pagesCUESTIONARIO 3 MicrobiologiaYohana SaucedoNo ratings yet
- Conceptos de VirologíaDocument8 pagesConceptos de VirologíaJoseNo ratings yet
- Silabo Microbiología Ambiental PDFDocument13 pagesSilabo Microbiología Ambiental PDFCristian DamianNo ratings yet
- Póster .pdf-PDFA PDFDocument1 pagePóster .pdf-PDFA PDFEvy Joy Portilla DiazNo ratings yet
- Importancia de La MicrobiologíaDocument1 pageImportancia de La MicrobiologíaRhuly Agustín VázquezNo ratings yet
- A Grim orDocument67 pagesA Grim orMaria Jose MartinNo ratings yet
- Trabajo de InvestigacionDocument11 pagesTrabajo de InvestigacionEliza Calle MoralesNo ratings yet
- Consulta Flagelo BacterianoDocument2 pagesConsulta Flagelo BacterianoMiriam Medina100% (2)
- Conservación de Cepas MicrobianasDocument11 pagesConservación de Cepas MicrobianasLuis Ferráez LindemannNo ratings yet
- Ficha R.solanacearumDocument9 pagesFicha R.solanacearumGabriel Segura C.No ratings yet
- Investifacion MicroDocument2 pagesInvestifacion MicroFernanda Quiroz chNo ratings yet
- Evidencia 2 - Diversidad BiologicaDocument26 pagesEvidencia 2 - Diversidad BiologicaLizette GalarzaNo ratings yet
- Kumar, 2017.en - EsDocument18 pagesKumar, 2017.en - Esazalia flor de frioNo ratings yet
- Estructura BacterianaDocument7 pagesEstructura BacterianaMartha De La VegaNo ratings yet
- Clasificación de Los Desinfectantes y AntisépticosDocument12 pagesClasificación de Los Desinfectantes y AntisépticosAnonymous GUXjVMXUNo ratings yet
- Teoría Endosimbiótica Ensayo PDFDocument5 pagesTeoría Endosimbiótica Ensayo PDFAlberto CruzNo ratings yet
- Capitulo 7 y 8Document7 pagesCapitulo 7 y 8Otoqui HernandezNo ratings yet
- KlebsielaDocument32 pagesKlebsielaVania Ponce LopezNo ratings yet
- Análisis Microbiólgico de La Carne 9Document8 pagesAnálisis Microbiólgico de La Carne 9laydemezalopez0% (1)
- Ctbiol 2ºs IipDocument10 pagesCtbiol 2ºs IipmalcommmNo ratings yet
- 56 BIOLOGIA ExamenDocument25 pages56 BIOLOGIA ExamenDaniela De Alba Stevenson100% (2)
- Objetivo 2. Taxonomia de Los Seres VivosDocument7 pagesObjetivo 2. Taxonomia de Los Seres Vivosmusicart30008394100% (1)
- Informe EnterobacteriasDocument4 pagesInforme EnterobacteriasmltorresNo ratings yet