You might also like
- Cancer Refers To Any One of A Large Number of Diseases Characterized by The Development of Abnormal Cells That Divide Uncontrollably and Have The Ability To Infiltrate and Destroy Normal Body TissueDocument11 pagesCancer Refers To Any One of A Large Number of Diseases Characterized by The Development of Abnormal Cells That Divide Uncontrollably and Have The Ability To Infiltrate and Destroy Normal Body TissueThess Tecla Zerauc AzodnemNo ratings yet
- Mayors of NCRDocument3 pagesMayors of NCRThess Tecla Zerauc AzodnemNo ratings yet
- Blocks in Math CountingDocument1 pageBlocks in Math CountingThess Tecla Zerauc AzodnemNo ratings yet
- Portfolio inDocument31 pagesPortfolio inThess Tecla Zerauc AzodnemNo ratings yet
- FluDocument4 pagesFluThess Tecla Zerauc AzodnemNo ratings yet
- Third Periodic Reivewer Science VIDocument5 pagesThird Periodic Reivewer Science VIThess Tecla Zerauc AzodnemNo ratings yet
- Jings Tutorial Center - Docx KinderDocument4 pagesJings Tutorial Center - Docx KinderThess Tecla Zerauc AzodnemNo ratings yet
- Envi CodeDocument2 pagesEnvi CodeThess Tecla Zerauc AzodnemNo ratings yet
- NameDocument1 pageNameThess Tecla Zerauc AzodnemNo ratings yet
- FluDocument9 pagesFluThess Tecla Zerauc AzodnemNo ratings yet
- FluDocument4 pagesFluThess Tecla Zerauc AzodnemNo ratings yet
- FluDocument4 pagesFluThess Tecla Zerauc AzodnemNo ratings yet
- FluDocument4 pagesFluThess Tecla Zerauc AzodnemNo ratings yet
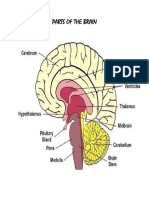
- Organs of The Human BodyDocument8 pagesOrgans of The Human BodyThess Tecla Zerauc AzodnemNo ratings yet
- Kinds of Musical InstrumentsDocument2 pagesKinds of Musical InstrumentsThess Tecla Zerauc AzodnemNo ratings yet
- Unders Tandin GIN Passin G Requir EmentsDocument4 pagesUnders Tandin GIN Passin G Requir EmentsThess Tecla Zerauc AzodnemNo ratings yet
- QC Environment CodeDocument2 pagesQC Environment CodeThess Tecla Zerauc Azodnem100% (4)
- LiverDocument2 pagesLiverThess Tecla Zerauc AzodnemNo ratings yet
- 1Document4 pages1Thess Tecla Zerauc AzodnemNo ratings yet
- Things To Do in LagunaDocument4 pagesThings To Do in LagunaThess Tecla Zerauc AzodnemNo ratings yet
- "God Will Protect You: "Pray and Don't Lose Heart."Document1 page"God Will Protect You: "Pray and Don't Lose Heart."Thess Tecla Zerauc AzodnemNo ratings yet
- The HumanDocument1 pageThe HumanThess Tecla Zerauc AzodnemNo ratings yet
- Refrigeration Tools Are Used in Performing Preventive Maintenance and Repair On Air ConditionersDocument5 pagesRefrigeration Tools Are Used in Performing Preventive Maintenance and Repair On Air ConditionersThess Tecla Zerauc Azodnem83% (6)
- Legend of The Christmas SpiderDocument4 pagesLegend of The Christmas SpiderThess Tecla Zerauc AzodnemNo ratings yet
- Si Malakas at Si MagandaDocument6 pagesSi Malakas at Si MagandaThess Tecla Zerauc Azodnem100% (2)
- Emergency and Hotline NumberDocument1 pageEmergency and Hotline NumberThess Tecla Zerauc AzodnemNo ratings yet
- Igneous RocksDocument3 pagesIgneous RocksThess Tecla Zerauc AzodnemNo ratings yet
- Arts and Crafts of LuzonDocument2 pagesArts and Crafts of LuzonThess Tecla Zerauc Azodnem100% (6)
- Part of Speech Function or "Job" Example Words Example SentencesDocument2 pagesPart of Speech Function or "Job" Example Words Example SentencesThess Tecla Zerauc AzodnemNo ratings yet
- Tess Tess Tess: Peanut Peanut PeanutDocument1 pageTess Tess Tess: Peanut Peanut PeanutThess Tecla Zerauc AzodnemNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Parkinson's DiseaseDocument13 pagesParkinson's DiseaseStella WatkinsNo ratings yet
- Chakras and The Endocrine SystemDocument11 pagesChakras and The Endocrine SystemNalini Muniyendi100% (2)
- NeuropelveologyDocument77 pagesNeuropelveologySantiago Tomas QuinterosNo ratings yet
- My Selective MutismDocument18 pagesMy Selective MutismMichelle DaquelNo ratings yet
- Neuromuscular 2 2010 With AnswersDocument13 pagesNeuromuscular 2 2010 With AnswersWenzy CruzNo ratings yet
- Intracerebral HemorrhageDocument12 pagesIntracerebral HemorrhagevizzieNo ratings yet
- NCP Multiple SclerosisDocument2 pagesNCP Multiple SclerosisJaylord Verazon100% (2)
- Pain ManagementDocument17 pagesPain Managementa bNo ratings yet
- Nim 2.0 System and Et Tube PlacementDocument1 pageNim 2.0 System and Et Tube PlacementShyam PillaiNo ratings yet
- Functional AssessmentDocument11 pagesFunctional AssessmentXlian Myzter YosaNo ratings yet
- Essay Learning StylesDocument7 pagesEssay Learning StylesNurulSyafiqahNo ratings yet
- Observable and Measurable Aspects of Human Behaviors: BehaviorismDocument5 pagesObservable and Measurable Aspects of Human Behaviors: BehaviorismGeethaanjali KathiwananNo ratings yet
- PL3102 NotesDocument8 pagesPL3102 NotesKatriel HeartsBigbangNo ratings yet
- Cyclic MeditationDocument8 pagesCyclic MeditationSatadal GuptaNo ratings yet
- Neuropsychological Basis of Learning and MemoryDocument24 pagesNeuropsychological Basis of Learning and MemoryAnanyaNo ratings yet
- Physiology Essay 11: Sympathetic Nervous System in Stress ResponseDocument2 pagesPhysiology Essay 11: Sympathetic Nervous System in Stress ResponseNektarios TsakalosNo ratings yet
- WHO Analgesic LadderDocument2 pagesWHO Analgesic LadderAndi PakdheeNo ratings yet
- Implantable Stimulators For Neuromuscular ControlDocument28 pagesImplantable Stimulators For Neuromuscular ControlLaurence Garcia LaurenteNo ratings yet
- Psychiatrics McqsDocument20 pagesPsychiatrics McqsHirwaNo ratings yet
- Mama Points ExcerptDocument16 pagesMama Points Excerptbromleydave100% (2)
- Lumbar Disc HerniationDocument4 pagesLumbar Disc HerniationChester NicoleNo ratings yet
- The Dyscalculia AssessmentDocument18 pagesThe Dyscalculia AssessmentContinuum67% (9)
- Bassetti Claudio, Michel Billiard, Emmanuel Mignot Narcolepsy and Hypersomnia PDFDocument728 pagesBassetti Claudio, Michel Billiard, Emmanuel Mignot Narcolepsy and Hypersomnia PDFSandro VoltaNo ratings yet
- Cerebral Protection & NeuroresuscitationDocument19 pagesCerebral Protection & NeuroresuscitationImad El SadekNo ratings yet
- SLR Test Dan BragardDocument17 pagesSLR Test Dan BragardjunedyNo ratings yet
- The Nervous SystemDocument21 pagesThe Nervous Systemmisterbrowner@yahoo.comNo ratings yet
- 1105 Health 2Document36 pages1105 Health 2confessor of heartNo ratings yet
- Aunt Minnie Pediatric NeuroDocument15 pagesAunt Minnie Pediatric NeuroRommel OliverasNo ratings yet
- Functional Neurological Disorder New Subtypes and Shared Mechanisms - CLINICALKEY - DR RivasDocument14 pagesFunctional Neurological Disorder New Subtypes and Shared Mechanisms - CLINICALKEY - DR RivasFernando Pérez MuñozNo ratings yet
- Craniotomy Case StudyDocument14 pagesCraniotomy Case StudyHoney Semafranca PlatolonNo ratings yet