You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Behavioral Checklist PDFDocument7 pagesBehavioral Checklist PDFMartin Castañeda Salas100% (1)
- Brain Wave Vibration PDFDocument26 pagesBrain Wave Vibration PDFnikanor1100% (4)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Healing Power of Rainforest Herbs PDFDocument532 pagesThe Healing Power of Rainforest Herbs PDFRaluca Huzum100% (10)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Test 2Document15 pagesTest 2Ding DialyzerNo ratings yet
- Nursing Care Plan - UtiDocument2 pagesNursing Care Plan - Utisweet_karisma05100% (6)
- Low Back Pain, 2O12Document361 pagesLow Back Pain, 2O12Bán Kata100% (4)
- Skin Lesion DiagnosisDocument9 pagesSkin Lesion DiagnosisaequNo ratings yet
- InTech-The Pathogenesis of VitiligoDocument27 pagesInTech-The Pathogenesis of Vitiligorakunn92No ratings yet
- Facts 2016 PDFDocument2 pagesFacts 2016 PDFrakunn92No ratings yet
- S.porta Dan PatogenesisDocument4 pagesS.porta Dan Patogenesisrakunn92No ratings yet
- Vitiligo 1Document12 pagesVitiligo 1rakunn92No ratings yet
- Vitiligo EpidemDocument10 pagesVitiligo Epidemrakunn92No ratings yet
- Vitiligo 1Document12 pagesVitiligo 1rakunn92No ratings yet
- Eye Health Statistic June 2009Document4 pagesEye Health Statistic June 2009rakunn92No ratings yet
- KatarakDocument6 pagesKatarakrakunn92No ratings yet
- Belazo Alkali Sealer MSDSDocument4 pagesBelazo Alkali Sealer MSDSrumahsketchNo ratings yet
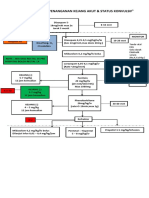
- Algoritma Penanganan Kejang AkutDocument1 pageAlgoritma Penanganan Kejang AkutEwa ClaudiaNo ratings yet
- Topik 1. Pengantar Epidemiologi Penyakit Menular - 2015Document47 pagesTopik 1. Pengantar Epidemiologi Penyakit Menular - 2015Arini Idza SafarinaNo ratings yet
- Blood Donation Application FormDocument2 pagesBlood Donation Application FormFadjar MulyaNo ratings yet
- Yellow Wallpaper Lit AnalysisDocument6 pagesYellow Wallpaper Lit Analysisapi-197908942No ratings yet
- CvadDocument11 pagesCvadNjideka A.No ratings yet
- An Approach To Common Urological Disorders PDFDocument45 pagesAn Approach To Common Urological Disorders PDFCrystel Tze JingNo ratings yet
- Brosur - Ageloc Youth enDocument20 pagesBrosur - Ageloc Youth enLidya ZhuangNo ratings yet
- Management of Side Effects and Complication in Medical AbortionDocument10 pagesManagement of Side Effects and Complication in Medical AbortionmariaNo ratings yet
- Tetracycline Drug Reporting-2Document21 pagesTetracycline Drug Reporting-2Shynne RPhNo ratings yet
- Strategias DE Prevención E Intervención DEL Burnout EN EL Ámbito EducativoDocument7 pagesStrategias DE Prevención E Intervención DEL Burnout EN EL Ámbito EducativoEuge López RuizNo ratings yet
- Pleural Puncture/thoracentesis: Pretty Laura ListianiDocument21 pagesPleural Puncture/thoracentesis: Pretty Laura ListianirastaNo ratings yet
- AcetaminophenDocument2 pagesAcetaminophenKimskiusNo ratings yet
- Pharmacovigilance Responsibilities of Medicine Sponsors: Australian Recommendations and RequirementsDocument44 pagesPharmacovigilance Responsibilities of Medicine Sponsors: Australian Recommendations and RequirementsResmy JoseNo ratings yet
- Navrekar - LWM PPT For Workshop 16 JulyDocument40 pagesNavrekar - LWM PPT For Workshop 16 JulyQudratullah ShaikNo ratings yet
- Blue Baby SyndromeDocument6 pagesBlue Baby SyndromeHerryanto AgustriadiNo ratings yet
- Manuscrip 2 PDFDocument41 pagesManuscrip 2 PDFayu purnamaNo ratings yet
- SystemOne 60 Series REMstar Auto User ManualDocument28 pagesSystemOne 60 Series REMstar Auto User Manualmanojsingh4allNo ratings yet
- Visionaire 2 3 Service Manual English PDFDocument47 pagesVisionaire 2 3 Service Manual English PDFQuarmina HesseNo ratings yet
- AINS and EyeDocument26 pagesAINS and EyeEcaterina Borovic-PavlovschiNo ratings yet
- Tinnitus Today September 1986 Vol 11, No 3Document8 pagesTinnitus Today September 1986 Vol 11, No 3American Tinnitus AssociationNo ratings yet
- Corpening Group Exercise January: RM Time Class Instruc. RM Time Class Instruc. RM Time Class InstrucDocument1 pageCorpening Group Exercise January: RM Time Class Instruc. RM Time Class Instruc. RM Time Class InstrucymcawncNo ratings yet
- Tacrine Induce Hepatotoksik PDFDocument9 pagesTacrine Induce Hepatotoksik PDFItamahYulaikhaNo ratings yet
- Case StudyDocument3 pagesCase StudyAvni DhingraNo ratings yet