Professional Documents
Culture Documents
Genética FCM-UNL (Sebenta) Ana Carlota Dias
Uploaded by
Miguelzip2000Original Description:
Original Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Genética FCM-UNL (Sebenta) Ana Carlota Dias
Uploaded by
Miguelzip2000Copyright:
Available Formats
-Actualizada-
2009-2010
Faculdade de Cincias Mdicas, Universidade Nova de Lisboa
Prefcio
Esta sebenta foi elaborada por Ana Carlota Dias, no 1
semestre do segundo ano do curso de Medicina da FCML no
ano lectivo 2009/2010 e surgiu da minha necessidade eminente
de ver a luz ao fundo do tnel nesta rea da gentica.
Assim, organizei-a de modo a cobrir todos os temas do
programa de gentica elaborado pelo professor Rueff para o
ano lectivo 2009/2010. Os poucos temas que no desenvolvi
foram os que achei totalmente chineses ou repetidos num ou
noutro ponto do programa.
Todo o texto desta sebenta foi literalmente copiado, quer da
sebenta de gentica nova que est neste momento em
circulao, quer dos slides das aulas tericas do professor e
ainda de livros e artigos, os quais esto presentes no ndice
bibliogrfico. Quero portanto salientar que nada do que aqui
est escrito da minha autoria.
Espero que vos seja til.
Gentica-Programa FCM / UNL Pgina 1 de 12
1
GENTICA
PROGRAMA DETALHADO
2009 / 2010
1. INTRODUO (3 horas).
1. A Gentica como rea cientfica multidisciplinar e em rpida evoluo.
Anlise histrica da evoluo da Gentica. De Miesher a Watson e Crick; de
Mendel a Morgan.
2. Gentica descritiva e Gentica mecanicista.
Incidncia, prevalncia e impacto em sade pblica da patologia gentica.
Taxonomia das doenas genticas e mecanismos de transmisso:
Monognicas.
Polignicas.
Multifactoriais.
Cromossmicas.
3. Dominncia completa e incompleta, codominncia, penetrncia e expressividade
de um gene.
Pleiotropia. Sries multiallicas. Alelos letais.
4. A histria clnica em Gentica e a rvore genealgica.
2. ANATOMIA E FISIOLOGIA DO MATERIAL GENTICO (6 horas).
1. Estrutura do polinucletido e nucleoproteica.
DNA e RNA.
Regies repetitivas: minisatlites, microsatlites, CNVs
Histonas e protenas no-histnicas do nucleossoma.
Do nucletido ao cromossoma.
2. Material gentico, ciclo celular e ciclo biolgico.
Fases G1, S, G2 e M do ciclo celular.
Noo de ponto de restrio.
Gentica-Programa FCM / UNL Pgina 2 de 12
2
G1/G0 quiescncia.
Controlo da proliferao celular e cascatas de transduo de sinal.
Fase S e replicao semi-conservativa e semi-descontnua do DNA.
RNA iniciador e fragmentos de Okasaky.
Protenas estabilizadoras da cadeia simples, helicases e topoisomerases 1 e 2.
O problema do fim da replicao: telomerase.
Fase M.
Condensao de cromatina.
Fuso acromtico e protenas contrcteis do citoplasma.
Mitose.
Recombinao mittica e troca de cromtides irms (SCEs).
O ciclo biolgico e a reduo meitica da ploidia.
Meiose.
Quiasmata e crossing-over.
Converso e complementao gnica.
3. Genes e expresso do material gentico
Transcrio.
RNA polimerases I, II e III.
Locais de iniciao.
Elementos cis e elementos trans.
Locais de ligao TATA, GC, CAAT e factores de transcrio.
Formao do cap e poli-adenilao.
Intres e exes.
snRNAs, spliceossoma e splicing dos RNAs. Splicing alternativo.
Edio do RNA.
RNAs no codificantes (ncRNA):
microRNAs e o mecanismo de interferncia de RNA. Ligao aos 3UTR
Rede de regulao mediada pelos ncRNAs.
Gentica-Programa FCM / UNL Pgina 3 de 12
3
Cdigo gentico.
Degenerescncia do cdigo gentico.
A quase universalidade do cdigo gentico.
Cdigo gentico mitocondrial.
Traduo.
Estrutura dos ribossomas, RNAs e protenas ribossomais
Activao dos aminocidos.
Amino-acil-tRNA sintetases.
Iniciao.
Elongao.
Peptidil-transferase e translocao.
Terminao.
Pptido sinal e topognese.
Regies no traduzidas 5UTR e 3UTR.
4. Regulao da expresso gentica.
Nveis de regulao da expresso gentica:
Regulao transcripcional
Regulao no processamento de mRNA e modificaes ps-transcripcionais.
Factores de transcrio, promotores, enhancers e silencers.
Regulao ps-transcrio.
Regulao e organizao da cromatina.
5. Gentica de Populaes
A uniformidade e a disjuno mendelianas dos caracteres.
Dominncia e recessividade.
Conceito de locus e de alelo.
Heterogeneidade gentica e allica.
Gentica-Programa FCM / UNL Pgina 4 de 12
4
A segregao mendeliana dos caracteres
Recombinao por crossing-over e o fenmeno de linkage.
Transmisso matrilinear do DNA mitocondrial.
Gentica formal.
Frequncia gnica, frequncia genotpica.
Equilbrio de Hardy-Weinberg.
Bialelia e poli-alelia.
Polimorfismos.
Isolados genticos.
Panmixia e endogamia.
Efeito fundador.
Conceito de bottleneck. Expanso e migrao.
Migraes populacionais de alelos.
Desequilbrio de linkage e sua variao a nvel populacional.
HapMap e blocos haplotpicos
Deriva gentica.
Fitness e distribuio geogrfica.
Incidncia e prevalncia de doenas genticas e fitness.
3. DIVERSIDADE E PATOLOGIA (6 horas).
1. Mecanismos e consequncias de mutao
Mecanismos de mutao e classificao de mutaes.
Classificao das mutaes:
Mutaes pontuais, deleces, inseres, duplicaes, expanso de
tripletos. Conceito de pr-mutao (expanses instveis de tripletos).
Mecanismos de mutao.
Tautomeria de bases.
Erros de proof reading e emparelhamentos errados.
Deslizamento da DNA polimerase em regies repetitivas.
Gentica-Programa FCM / UNL Pgina 5 de 12
5
Desaminao espontnea da 5-metil citosina e hipermutabilidade das regies
CpG.
Mutaes induzidas por mutagneos fsicos e qumicos.
Radiao ionizante e UV.
Agentes qumicos intercalantes e anlogos de bases.
Agentes qumicos alquilantes e arilantes. Aductos de DNA.
Metabolizao de pr-mutagneos (Metabolismo de fase 1 e de fase 2).
A super famlia gnica dos citocromos P450 (genes CYP).
Inducibilidade e susceptibilidade a cancro.
Significado e consequncias de mutaes.
Em regies extra-gnicas: annimas (SNPs, VNTR e STR).
Em regies codificantes.
Silenciosas, missense, nonsense, frameshift, de splicing.
Em regies reguladoras.
Consequncias patolgicas em cada tipo de mutao.
Tipos de mutao mais frequentes em patologia humana.
Impacto em patologia gentica da taxa de neo-mutao humana.
2. Mecanismos protectores da fidelidade informativa.
Sistemas enzimticos de reparao do DNA.
Reparao pr-replicativa pelas DNA-polimerases.
Reparao ps-replicativa por recombinao.
Reparao por exciso
Glicosilases e endonucleases de locais apurnicos.
NER, BER, MMR. NHEJ
Sndromes de dfice de reparao e fragilidade cromossmica.
Anemia de Fanconi.
Ataxia-telangiectasia.
Sndrome de Lynch e dfice de reparao de emparelhamentos errneos MMR
Gentica-Programa FCM / UNL Pgina 6 de 12
6
Xeroderma pigmentosum.
Sndrome de Cockayne.
Trico-tiodistrofia.
Sndrome de Bloom e o dfice de DNA-ligase.
4. DA DIVERSIDADE PATOLOGIA - EXPRESSO CLNICA E MECANISMOS DE
DOENA GENTICA (8 horas).
1. Patologia gentica monognica.
Fibrose qustica.
O gene CFTR, transporte trans-membranar de cloreto e a clnica da fibrose
qustica.
A mutao F508 e as outras mutaes do gene CFTR. Heterozigotos
compostos.
A migrao e expanso europeia de alelos mutantes de CFTR.
Classificao das mutaes no gene CFTR. Do laboratrio clnica.
Glucosuria renal. Mutao no gene SLC5A2. Mutaes privativas
Sndrome de Antley-Bixler. Mutao no gene POR.
Hemoglobinopatias.
Hemoglobina Lepore e crossing-over desigual.
Talassmias-deleco, represso da transcrio ou traduo e anomalias de
splicing.
Drepanocitose e haplotipos de restrio.
Origem e distribuio das mutaes nas hemoglobinopatias e gentica de
populaes.
Quadro clnico das hemoglobinopatias.
2. Patologia gentica monognica autossmica dominante.
Anlise de mecanismos de expresso clnica autossmica dominante.
Alteraes nos colagneos.
Osteognese imperfeita e sndrome de Elhers-Danlos dominante.
Gentica-Programa FCM / UNL Pgina 7 de 12
7
A mutao da transtirretina e polineuropatia amiloidtica familiar (PAF).
O sndrome de Marfan e a fibrilhina.
Coreia de Huntington.
Neurofibromatose de von Recklinghausen.
A cardio-miopatia hipertrfica familiar e heterogeneidade de locus e allica.
Doena poliqustica renal.
3. Genopatias metablicas.
Doenas lisossomais - mucopolissacaridoses e esfingolipidoses.
A doena de Tay-Sachs e a endogamia.
Sndromes de Hurler, Hunter e I cell disease - anlise dos mecanismos
etiopatognicos.
Terapia gnica nas doenas de Fabry, de Gaucher e no sndrome de Hurler.
As vias metablicas da fenilalanina e tirosina - fenilcetonria, alcaptonria e
albinismo.
Deficincia de alfa-1-antitripsina.
As hiper-lipoproteinmias familiares - anlise dos dfices de receptores de LDL.
Doenas peroxissomais:
Sndrome de Zellweger.
Adrenoleucodistrofia.
Doena de Refsum - da mutao pontual ao dfice de sinais de topognese.
4. Mosaicismo da expresso ligada ao cromossoma X.
Mecanismos de inactivao do cromossoma X.
Hemoflia e sndrome de Lesh-Nyan.
A distrofina e a distrofia muscular de Duchenne.
Deficincia em glucose-6-fosfato desidrogenase.
5. Hereditariedade no convencional.
Dissomias uniparentais.
Gentica-Programa FCM / UNL Pgina 8 de 12
8
Imprinting genmico: sndromes de Prader-Willi e de Angelman.
Expanso de tripletos como mecanismo de doena e sndromes de expanso de
tripletos.
X-frgil.
Coreia de Huntington e outras doenas de poliglutamina.
Ataxia de Friedreich
Distrofia miotnica.
Doena de Machado-Joseph.
6. Patologia citogentica.
Citogentica clnica.
Polimorfismos e abortamentos de repetio.
Metodologias de bandeamento.
Aneuploidias de expresso fenotpica patolgica - as trissomias 13, 18 e 21.
Deleces e microdeleces.
Sndrome do grito do gato e variabilidade das extenses de deleco de 5p.
Sndrome de DiGeorge.
Microdeleco do cromossoma Y e infertilidade.
Deleces em 1p e 19q em oligodendrogliomas, significado prognstico e
teraputico.
Sndromes de genes contguos (DiGeorge, Smith-Magenis, Williams).
Translocaes equilibradas, transmisso e clculo de risco.
Inverses pericntricas e paracntricas.
Cromossomas em anel e isocromossomas.
Outras anomalias estruturais (dicntricos, fragmentos acntricos, entre outros).
Mosaicismo citogentico.
Fragilidade cromossmica e mapa de locais frgeis.
Da clnica citogentica e anlise molecular do sndrome do X frgil.
Gentica-Programa FCM / UNL Pgina 9 de 12
9
7. Patologia multifactorial.
Situaes multifactoriais.
Identificao de genes de susceptibilidade em doenas multifactoriais.
Estudos de associao de caso-controlo e estudos familiares.
Marcadores annimos e desequilbrio de linkage.
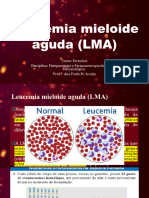
5. A GENTICA DO CANCRO (3 horas).
1. Mutao somtica e cancro.
Oncogenes e genes supressores de tumores.
O carcter multifsico e multifactorial da cancerignese. Hipteses gentica e
epigentica.
Oncogene addiction. Passenger and Driver mutations.
Leucemia mielide crnica e tirosina-quinases. Receptores de transporte e
resistncia teraputica.
2. Sndromes de cancro familiar.
Mutaes nos genes BRCA1 e BRCA2 no cancro da mama familiar.
Mutao no gene RB e retinoblastoma hereditrio por perda de heterozigotia (LOH).
A polipose adenomatosa familiar (FAP) e o sndrome de Lynch (HNPCC).
P53 e sndrome de Li-Fraumeni.
Haploinsuficincia e dominncia negativa (p53).
MEN1 e MEN2 (multiple endocrine neoplasia type 1 and type 2).
Alteraes cromossmicas e cancro
Deleco no cromossoma 11 e tumor de Wilms.
Deleco no cromossoma 13 e retinoblastoma.
Citogentica de clulas neoplsicas.
A actividade enzimtica da telomerase e o estudo de neoplasias.
Gentica-Programa FCM / UNL Pgina 10 de 12
10
6. A GENTICA MOLECULAR HUMANA - REAS DE INTERVENO (6 horas).
1. Testes genticos predictivos.
Diagnstico pr-implantatrio.
Diagnstico pr-natal.
Diagnstico pr-sintomtico.
Aconselhamento gentico, avaliao de risco e preveno da recorrncia.
2. Rastreios e diagnstico precoce (fenilcetonuria e hipotiridismo).
3. A Medicina Molecular.
A farmacogenmica e a oncogenmica.
A identificao de novos alvos teraputicos.
A engenharia de tecidos e a regenerao celular.
Lisboa, 28 de Setembro de 2009
O regente,
Jos Rueff
(Prof. Catedrtico)
Gentica-Programa FCM / UNL Pgina 11 de 12
11
GENTICA
Ano lectivo 2009 - 2010
Livros sugeridos
Principais livros de estudo:
Vogel and Motulsky's Human Genetics. Problems and Approaches
Speicher, Michael; Antonarakis, Stylianos E.; Motulsky, Arno G. (Eds.)
Springer Verlag, 4th ed., 2010.
Strachan T. and Read A.P. Human Molecular Genetics,
3
rd
edition. Garland Science, Taylor & Francis Group, 2004.
Livros de estudo adicionais:
Gardner, R.J.M. e Sutherland, G.R. Chromosome Abnormalities and Genetic
Counselling, 3
rd
edition. Oxford University Press, 2003.
Nussbaum R.L., McInnes R.R. e Willard H.F. Thompson & Thompson Genetics in
Medicine, 7
th
edition. Saunders, 2007.
Korf, B.R. Human Genetics and Genomics, 3
rd
edition. Blackwell, 2007.
Jorde L.B., Carey J.C., Bamshad M.J. e White R.L. Medical Genetics 3
rd
edition.
Mosby, 2005.
Lewis R. Human Genetics: Concepts and Applications. 8
th
edition. McGraw-Hill
Higher Education. 2008.
Turnpenny, P. e Ellard S. Emery's Elements of Medical Genetics. 13
th
edition.
Churchill Livingstone, 2007.
Passarge E. Color Atlas of Genetics. 3
rd
edition. Thieme Medical Publishers, 2007.
Gentica-Programa FCM / UNL Pgina 12 de 12
12
Regateiro F.J. Manual de Gentica Mdica. Imprensa da Universidade, Coimbra, 2003.
Livros para consulta:
Emery, A. and Rimoin, D. Principles and Practice of Medical Genetics" - 3 vols. 5
th
edition. Churchill Livingstone, 2007. (principalmente o 1 volume)
Trent R. Molecular Medicine" 3
rd
edition. Academic Press, 2005.
Encyclopedia of the Human Genome 5 vols. John Wiley & Sons, Ltd. 2003.
The Metabolic and Molecular Bases of Inherited Disease. 3 vols. Edited by Scriver,
C. R., Beaudet, A. L., Sly, W. S., Valle, D., Childs, B., Kinzler, K. W., and Vogelstein, B.,
eds., 8th ed., McGraw-Hill, New-York, 2001
Watson, J.D. A Passion for DNA; Genes, Genomes, and Society. Cold Spring Harbor
Laboratory Press, 2000.
Sebenta de Gentica
FCML
1
INTRODUO
1. A Gentica como rea multidisciplinar e em rpida evoluo_____
Anlise histrica da evoluo da Gentica. De Miesher a Watson e Crick; de
Mendel a Morgan
Sc. XIX
- Leis de Mendel
- Descoberta do DNA: Friedrich Miescher
Sc. XX Sculo do gene
- Re-descoberta das leis de Mendel
- Baptismo da Gentica (Bateson, 1906)
- Erros inatos de metabolismo (Garrod, 1909)*
- DNA transformante (Avery, McLeod, 1994)
- Complementaridade de bases (Erwin Chargaff)
- Estrutura do DNA (Watson e Crick)
- Citogentica e citogentica mdica (tjio e Levan; Lejeune)
- Lysenko (1887-1943) contraria as leis de Mendel.
- Thomas Hunt Morgan (1933)- prmio nobel por provar que os cromossomas so portadores
de genes
- Avery e MacLeod (1944) descobriram que o DNA determina as caractersticas genticas.
- Chargaff e Francis Crick complementaridade de bases de DNA.
- Wayson e Crick (e tambm, Rosalind Franklin) estrutura do DNA em dupla hlice.
Sc. XXI- Sculo do genoma
- Mapeamento gnico
- Sequenciao de DNA
- Programa do Genoma Humano
- Genmica, trascriptmicas, protemica, metabolmica, interactmica.
Prmio Nobel 2009 Telmeros (ver pg. #)
2. Gentica descritivia e mecanicista____________________________
A gentica descritiva a gentica da observao dos fenmenos. Regista e descreve as
caractersticas fenotpicas de indivduos e populaes. morfolgica e responde questo
como determinado indivduo em relao a uma dada caracterstica. Utiliza a elaborao
de rvores genealgicas. A gentica mecanicista com base no genoma, explica as alteraes
Sebenta de Gentica
FCML
2
genotpicas. fisiolgica e responde questo como funciona. Tem primeiro conhecimento
do gene e s depois da sua expresso e efeito. A gentica mecanicista corresponde
gentica experimental, que recorre criao de modelos e ao uso de animais transgnicos,
nos quais, atravs da fertilizao in vitro, se introduz DNA alterado (humano ou de outra
espcie), obtendo-se, desta forma, um exemplar vivo de determinada alterao, que pode ser
manipulado.
Incidncia, prevalncia e impacto em sade pblica da patologia gentica
Taxonomia das doenas genticas e mecanismos de transmisso:
- Monognicas- doenas que resultam de mutaes em apenas um gene
- Polignicas- doenas que resultam da aco combinada de alelos de mais de um gene.
- Multifactoriais- doenas em que uma parte pertence aos genes que conferem determinada
susceptibilidade (predisposio) gentica mas apresentam, igualmente, uma parte ambiental,
sem a qual a doena no se expressa.
- Cromossmicas- doenas causadas pela alterao da estrutura/nmero dos cromossomas.
3. Algumas definies_____________________________________________
Penetrncia- trata-se de um fenmeno de tudo-ou-nada e aplica-se probabiliade de um
gene se expressar ou no, isto , de ter ou no expresso fenotpica. Quando a frequncia
de um fentipo inferior a 100%, ou seja, quando alguns indivduos que tm o gentipo no
o expressam, diz-se que o gene exibe uma penetrncia incompleta. Em termos estatsticos,
a percentagem dos indivduos com um determinado gentipo que so, de facto, afectados.
Sebenta de Gentica
FCML
3
Expressividade de um gene- corresponde ao modo de expresso do alelo, que pode ser
uniforme (ocorre quando um alelo expressa sempre um nico tipo de fentipo, de fcil
reconhecimento) ou varivel (quando a expresso do alelo resulta no aparecimento de vrios
padres de fentipos ou vrios graus de expresso).
Dominncia completa- neste caso um alelo capaz de suprimir a manifestao do outro
quando em heterozigotia prevalecendo no fentipo.
Dominncia incompleta situaes em que o fentipo dos indivduos heterozigsticos
intermedirio, em termos quantitativos, entre os fentipos dos dois homozigticos. Ocorre em
indivduos heterozigticos que apresentam fentipos intermdios entre os seus progenitores
de linhagens puras, isto acontece porque uma nica copia do gene funcional no ser
suficiente para assegurar o fentipo, em outras palavras a expresso gnica de um nico
gene no suficiente para produzir uma quantidade mnima de enzima, por exemplo.
Codominncia- ocorre quando ambos os alelos de um gene se expressam integralmente no
fentipo do heterozigoto. Ex: grupos sanguineos AB0
Hemizigotia- presena de um nico alelo no genoma, condio que se verifica, normalmente,
para a grande maioria dos loci do cromossoma X, nos indivduos do sexo masculino.
Corresponde tambm a condies anormais (ex. numa deleco), em que em vez de um
gentipo diplide, se encontra uma nica cpia de um alelo.
Pleiotropia- no pleitropismo, a mutao de um nico gene tem reflexos na expresso de
vrios fentipos, pelo facto de uma nica protena actuar em diversos orgos do corpo
humano. Um exemplo do efeito pleiotrpico observa-se na neurofibromatose tipo I (ver pgina
140). O reconhecimento do pleiotropismo pode permitir que, a partir de uma manifestao,
ainda que minor, pertencente ao espectro de efeitos conhecidos de uma mutao, seja
possvel a identificao de um portador dessa mutao ou se suspeite de anomalias internas
graves que exijam interveno mdica.
Alelos letais- alelos cuja manifestao fenotpica corresponde morte do indivduo, seja na
fase pr-natal ou ps-natal. Os alelos letais dominantes surgem de mutaes de um alelo
Fig.1: dominancia incompleta
Sebenta de Gentica
FCML
4
normal. Os alelos letais recessivos s resultam na morte do indivduo quando em
homozigotia.
Sries multiallicas= alelos mltiplos. Mais do que dois alelos por gene.
4. Histria clnica em gentica e a arvore genealgica_____________
Fig. 2: Smbolos utilizados na rvore genealgica
ANATOMIA E FISIOLOGIA DO MATERIAL GENTICO
1. Estrutura do polinucletido e nucleoproteica___________________
DNA e RNA
DNA: Os cromossomas presentes no ncleo de cada clula, tm como constituinte principal,
uma complexa molcula de peso molecular muito elevado, o cido desoxirribonucleico
DNA. Este constitui o suporte material dos caracteres hereditrios. Na estrutura da molcula
esto efectivamente inscritas, segundo um cdigo particular, todas as informaes que
definem um ser vivo. O DNA encontra-se essencialmente no ncleo e nas mitocndrias.
uma macromolcula polimrica linear, formada pela polimerizao de unidades elementares
os nucletidos. Cada nucletido formado atravs da ligao covalente de trs molculas:
- uma pentose, a 2- desoxirribose;
- uma base orgnica contendo azoto
- um grupo fosfato (cido fosfrico - H
2
PO
3
)
As bases azotadas so de dois tipos: purinas e pirimidinas. As purinas so compostas por
uma estrutura em anel duplo - adenina (A) e guanina (G). As pirimidinas- citosina (C) e timina
(T) so constitudas apenas por um anel.
DNA monocatenrio: os nucletidos so polimerizados em longas cadeias polinucleotdicas
lineares atravs de ligaes fosfodister 5- 3. Estas ligaes efectuam-se entre o grupo
fosfato do carbono-5 da pentose de um dos nucletidos e o grupo hidroxilo livre do carbono
3 da pentose do nucletido que o precede na cadeia polinucleotdica formando a estrutura
primria do DNA DNA monocatenrio.
Sebenta de Gentica
FCML
5
DNA bicatenrio: embora certas formas de DNA celular possam existir sob a forma de
estruturas de cadeia simples, a estrutura de DNA mais generalizada a dupla hlice do
modelo introduzido por Watson & Crick segundo o qual, o DNA constitudo pela reunio
(por pontes de hidrognio) de duas longas cadeias polinucleotdicas (anti-paralelas), cuja
estrutura helicoidal se assemelha a uma escada em espiral orientada para a direita. Esta
estrutura secundria do DNA, sob a forma de dupla hlice, designada por DNA bicatenrio.
Topologicamente, o DNA pode existir na forma relaxada (relaxed DNA) ou na forma
superenrolada (supercoiled DNA). O DNA pode assumir vrias conformaes e pode haver
interconverso de formas:
- A enrolado para a direita (+ longo e curto que B)
- B - enrolado para a direita (forma predominante. Modelo de Watson & Crick)
- Z enrolado para a esquerda (+ delgada e alongada que A e B); rico em G e C, que
podem formar zonas de regulao associadas a zonas de silenciamento. As bases
azotadas esto voltadas para o exterior. So pouco frequentes.
DNA Mitocondrial: ver pgina 35.
RNA: o RNA cido ribonucleico um polmero linear de nucletidos de purina e pirimidina,
unidos por ligaes fosfodister 5'-3 com uma estrutura semelhante do DNA
monocatenrio, sendo a principal diferena entre eles a pentose constituinte dos nucletidos,
que no DNA a 2-desoxirribose e no RNA a ribose. Os ribonucletidos presentes so de
adenina (A), citosina (C) e guanina (G), tal como no DNA, no entanto, no RNA, a timina
substituda pelo Uracilo (U). Tendo em conta que a molcula de RNA complementar apenas
de uma das cadeias de DNA, a soma das bases pricas no necessariamente igual soma
das bases pirimdicas. Qualquer que seja o tipo ou funo que desempenham, os RNA so
sempre sintetizados por cpia de fragmentos especficos bem delimitados do DNA que
constitui o genoma de cada espcie, obedecendo a sua sntese ao princpio da
complementaridade de bases, atravs de um processo de transcrio. Neste processo, a
sequncia de bases de uma das cadeias de DNA (template strand) enzimaticamente
copiada sob a forma de uma cadeia simples, pelo que esta resulta semelhante outra cadeia
de DNA (coding strand).
RNA mensageiro (mRNA): o mRNA transporta a informao necessria sntese das
protenas, do ncleo ao local de sntese, isto , aos ribossomas (livres no citoplasma, nas
mitocndrias, associados ao retculo endoplasmtico). O mRNA sintetizado no ncleo
(podendo tambm ser sintetizado nas mitocndrias a partir do DNA mitocondrial), durante o
Sebenta de Gentica
FCML
6
processo de transcrio. O mRNA eucaritico sempre monocistrnico, ou seja, contm
informao para uma nica cadeia polipeptdica. Depois de ser transcrito e de sofrer o
processo de maturao, o mRNA passa aos ribossomas onde actua como molde indicador da
sequncia de aminocidos de uma cadeia polipeptdica a sintetizar. Tripletos de nucletidos,
ao longo da cadeia de mRNA, os codes, determinam, no processo de traduo, de um modo
especfico, a sequncia de aminocidos do polipptido. Portanto, a cada 3 bases do mRNA
corresponde um aminocido da protena. A esta correspondncia foi atribudo a designao
de cdigo gentico.
RNA ribossmico (rRNA): o rRNA constitui a maior parte do RNA celular, formando at 80%
do RNA total da clula. Nele est contido a informao para a formao das duas
subunidades dos ribossomas. Os ribossomas so organelos citoplasmticos resultantes da
associao do rRNA com protenas (complexos ribonucleiproteicos) e cuja funo principal
a sntese proteica a partir do mRNA.
RNA transferncia (tRNA): O tRNA pode constituir at 15% do RNA total da clula. As suas
molculas, relativamente pequenas, funcionam como transportadores de aminocidos e
interagem especificamente com o rRNA e mRNA no processo de traduo. A sua funo ,
primeiramente, activar os aminocidos de forma a que o estabelecimento da ligao peptdica
seja energeticamente favorvel. Seguidamente, o aminocido transferido para o
polirribossoma, onde incorporado no polipptido que est a ser sintetizado. A segunda
funo do tRNA o reconhecimento dos codes de mRNA, atravs do seu prprio anticodo,
que complementar ao codo do mRNA correspondente ao aminocido que transporta, por
forma a que este ltimo seja incorporado correctamente. H, portanto, pelo menos uma
molcula de tRNA para cada aminocido, podendo contudo, haver mais (tRNAs especficos
para o mesmo aminocido designam-se de isoaceitadores). A molcula de tRNA consiste
numa cadeia polinucleotdica formada por 65 a 110 mononucletidos e caracteriza-se por
conter, alm das bases comuns, uma proporo relativamente elevada (que pode atingir 10%
do total) de bases azotadas modificadas (bases que sofreram metilao ou dimetilao) e
ainda timina, constituinte normal do DNA, mas que no aparece em nenhuma outra espcie
de RNA. A metilao tambm diminui o carcter hidrofbico de algumas regies de tRNA, o
que pode ser relevante na interaco com sintetases e protenas ribossomais. O tRNA
apresenta a extremidade 5' fosforilada. A extremidade 3' constituda por uma sequncia de
bases caracterstica-CCA ( ao grupo 3'-OH do resduo de adenina que se liga um
aminocido especfico atravs de uma ligao ster com o grupo carboxilo). Embora cada
tRNA difira dos outros na sua estrutura primria, a estrutura secundria e terciria de todos
eles muito semelhante, possuindo uma conformao caracterstica em "folha de trevo"
Sebenta de Gentica
FCML
7
quando observada bidimensionalmente, devido existncia de uma extensa
complementaridade dentro da cadeia polinucleotdica. Deste modo, coexistem regies onde
as bases emparelham de forma idntica ao que se passa na cadeia dupla do DNA - regies
complementares denominadas braos (stems) e zonas onde as bases no so
complementares, constituindo ansas (loops). Cada tRNA possui na sua molcula dois stios
dotados de especificidade, que so a extremidade 3'-OH, que se liga ao aminocido e um
tripleto anticodo que se localiza no brao que ocupa a posio diametralmente oposta e
que ir reconhecer o codo correspondente no mRNA, por complementaridade de bases.
Para alm do brao aceitador (transportador de a.a.) e do brao anticodo, distingue-se,
ainda, um brao D (caracterizado pela presena da base dihidro-uridina) e um brao TC
(que possui a sequncia T, pseudouridina, C). Os tRNA so transcritos sob a forma de
grandes molculas precursoras, frequentemente contendo a sequncia para a sntese de
mais do que um tRNA, sendo seguidamente sujeitos a um processamento nucleoltico
catalisado por uma classe especfica de ribonucleases, aps o que adquirem dimenses mais
reduzidas. Para alm disto, as molculas de tRNA precursoras contm um intro contendo 10
a 40 nucletidos prximo da regio correspondente ao brao anticodo, pelo que o
processamento inclui tambm a clivagem dos intres e o splicing adequado da regio do
anticodo, de forma a originar uma molcula funcional. O sistema enzimtico responsvel
pelo processamento nucleoltico tem aparentemente a capacidade de reconhecer no
somente a estrutura primria do tRNA mas tambm a sua conformao espacial, pelo que s
as molculas que possuam uma estrutura funcionalmente competente sero processadas. A
modificao posterior das molculas de tRNA inclui a alquilao de certos resduos
nucleotdicos, bem como a incorporao do tripleto de CCA caracterstico da extremidade 3.
Esta ltima modificao realizada j no citoplasma e ocorre regularmente, visto que a
degradao das extremidades moleculares ocorre no citoplasma a uma muito maior
velocidade do que a da prpria molcula.
Sebenta de Gentica
FCML
8
Regies repetitivas: minisatlites, microsatlites, CNVs
DNA satlite: zonas repetidas bastante grandes (sequncias de 100Kb a vrios Mb)
que constituem a heterocromatina e os centrmeros. As regies extra podem levar a
situao de patologia.
Minisatlites: o conjunto global que ocupam no genoma mais pequeno que o DNA
satlite. Importantes para a identificao de aprtipos. Os minisatlites constituem os
telmeros e vrias regies cromossmicas.
Microssatlites: constitudas por repeties montonas (2 ou 3 bases); repeties
frequentes de Cs e As. Numa famlia podemos saber quem possui determinados
microsatlites atravs da anlise de linkage.
CNVs: copy-number variant. Os CNVs podem ocorrer por: repeties multialelicas;
arranjo complexo; inverso; duplicao. A Variao mdia de CNVs entre 12 a 14%
(a variao no muito elevada, no sendo variantes do genoma muito polimrficos
na populao). Os CNVs representam cerca de 17,0% da variao total da expresso
gnica (sendo que os restantes 83% se devem aos SNPs-single nucleotide
polimorfism). A sequenciao no identifica CNV.
Histonas e protenas no-histnicas do nucleossoma
Histonas: as histonas constituem uma famlia de protenas bsicas e heterognias, que
participam, juntamente com o DNA, na formao dos nucleossomas a unidade estrutural
bsica da cromatina. H 5 tipos principais de histonas designadas H1, H2A, H2B, H3 e H4,
diferindo principalmente pelo seu teor em arginina e lisina.
- Histona H1 - a que apresenta ligaes mais fracas cromatina, sendo facilmente
removida com uma soluo salina, aps o que a cromatina se torna solvel. Apresenta
um peso molecular duas vezes superior ao das outras histonas e caracteriza-se pelo seu
elevado contedo em aminocidos bsicos, particularmente lisina. responsvel pela
integridade e adeso dos diferentes nucleossomas.
- Histonas Nucleossmicas - H2A, H2B, H3 e H4 - estas protenas possuem uma estrutura
muito semelhante, que permaneceu constante entre as espcies ao longo da evoluo.
So particularmente ricas em arginina (H3 e H4) e em lisina (H2A e H2B) So
responsveis pela constituio dos nucleossomas e cada uma delas participa com duas
Sebenta de Gentica
FCML
9
subunidades, agrupando-se num octmero e formando o chamado core proteico do
nucleossoma ou complexo histnico. Em volta deste enrola-se a dupla hlice de DNA,
descrevendo uma volta e . Apresentam uma extremidade bsica que permite a ligao
ao DNA e uma extremidade hidrofbica que permite a associao entre elas.
Fig. 3: nucleossoma e distribuio das histonas
Protenas cidas ou no histnicas (NHP)
As protenas no histnicas constituem uma famlia muito heterognia com elevada
especificidade tecidular, que podem ser agrupadas em dois grupos distintos:
- Protenas constitutivas a actina, essencial para o mecanismo de formao do fuso
acromtico na metafase; molculas proteicas reguladoras, sensveis aco de
hormonas; protenas constituintes da arquitectura do prprio ncleo.
- Enzimas todas as enzimas necessrias aos processos de replicao, transcrio,
reparao e maturao do mRNA.
Protenas HMG (high motility group proteins): Grupo de protenas no histnicas cujo nome
deriva do facto de estas migrarem mais rapidamente em electroforese em gel uma vez que
so muito pequenas e apresentam uma elevada densidade de cargas. Estas protenas so
uns dos elementos que promovem a transio da cromatina da forma compacta para a forma
descondensada, transcricionalmente activa, pois interagem preferencialmente com a
cromatina na forma de fibra de 10nm.
Do nucletido ao cromossoma
Nveis de organizao estrutural do DNA: a interaco das histonas com o DNA resulta, como
j falmos, numa estrutura denominada nucleossoma. Os nucleossomas encontram-se
ligados entre si por DNA linker. Cada nucleossoma uma estrutura discide, constituda por
um ncleo central histnico (um octmero formado por duas histonas de cada um dos tipos
Sebenta de Gentica
FCML
10
H3, H4, H2A e H2B) com cerca de 10 nm de dimetro, em torno do qual o DNA se dispe em
super-hlice, em duas voltas orientadas para a esquerda, estabilizadas pela histona H1. A
organizao do DNA em polinucleossomas parece ser mediada por uma protena no
histnica polianinica, a nucleoplasmina. Esta protena no tem a capacidade de se ligar ao
DNA nem cromatina, mas estabelece uma interaco reversvel com as histonas, de modo
que a interaco destas com o DNA se torna muito mais especfica, sendo libertada da
estrutura logo aps a formao do nucleossoma. Embora os nucleossomas se encontrem
periodicamente posicionados ao longo da estrutura polinucleossomal, a sua distribuio no
aleatria com respeito sequncia de bases com que interagem. Com efeito, os
nucleossomas parecem exibir preferncia para certas sequncias de DNA, fenmeno que
designado de phasing. Este parece estar relacionado com a flexibilidade relativa dos vrios
tipos de sequncia e com a consequente maior ou menor facilidade de acomodao do DNA
em torno das histonas. Como as sequncias ricas em A-T so mais fceis de comprimir do
que as ricas em C-G, cada octmero de histonas tende a posicionar-se de modo a maximizar
a interaco com sequncias ricas em A-T. Uma outra caracterstica dos nucleossomas a
sua mobilidade. A maioria dos nucleossomas tem a capacidade de se posicionar mais a
montante ou mais a jusante numa distncia de 10 bp, o que facilita o acesso das DNA
polimerases e outras enzimas a essas sequncias.
Nos nucleossomas, o DNA condensado para cerca de 1/10 do seu comprimento aparente,
dando origem chamada fibra de 10nm, correspondente ao dimetro do nucleossoma. Neste
nvel, o DNA e as histonas constituem o nucleofilamento.
Estes nucleofilamentos vo ainda ser compactados em estruturas helicoidais de cromatina,
designadas de solenides (ver figura 4), que ao microscpio electrnico aparecem como
fibras grossas de 34 nm de dimetro, obtendo-se um encurtamento de 50%. Esta fibra,
designada por fibra de 30 nm, parece ser a unidade fundamental da cromatina. Cada volta do
solenide contm cerca de 6 nucleossomas.
Fig. 4: Estrutura do Solenide
Sebenta de Gentica
FCML
11
A histona H1 parece desempenhar um papel importante na compactao da fibra de 30 nm,
na medida em que tm a capacidade de se ligar cooperativamente entre si, aproximando os
nucleossomas uns dos outros. As fibras de 30 nm formam-se selectivamente em regies
especficas do DNA, caracterizadas pela ausncia de ligao a NHP especficas de
determinadas sequncias. Pensa-se que a presena destas protenas est relacionada com o
estado transcripcional das regies envolvidas. Os niveis de organizao desde a estrutura em
solenide ao cromossoma em metafase ainda no esto totalmente compreendidos
considerando-se porm a existncia um estado de maior compactao, que corresponde ao
super-solenide, cujas fibras apresentam um dimetro de 200 nm.
Eucromatina: ou cromatina activa, encontra-se descondensada e corresponde s regies do
genoma transcricionalmente activas. Contm normalmente uma maior densidade de regies
codificantes e a primeira a ser a replicada na fase S.
Heterocromatina: ou cromatina inactiva constitui cerca de 80% do DNA nuclear e apresenta-
se condensada, sendo bem visvel no ncleo em interfase, onde se evidencia como regies
densas e fortemente coradas, os crommeros.
- Heterocromatina facultativa: corresponde eucromatina, em condio inactiva para
a transcrio (ex. cromossoma X sujeito lionizao)
- Heterocromatina constitutiva: encontra-se junto dos centrmeros de todos os
cromossomas, no brao longo do cromossoma Y e nos satlites dos cromossomas
acrocntricos. Corresponde ao DNA satlite.
Territrios cromossmicos no ncleo:
- Os cromossomas ocupam no ncleo territrios no aleatrios.
- Os territrios cromossmicos no so entidades slidas, mas so permeadas por canais.
- Alteraes relativas de regies genmicas podem ser funcionalmente relevantes para
activao, represso e para translocaes.
- Pode determinar translocaes recprocas preferenciais entre cromossomas.
Genoma:
% de DNA codante total do genoma 1,5%
% de DNA codante/gene 5%
Possui alguns genes ortlogos (1,5%)
Possui graus de repetio muito grandes.
Sebenta de Gentica
FCML
12
Fig.5: % relativa de genes por funo
- Genes ortlogos genes de espcies diferentes que evoluram de um gene ancestral
comum por especiao. Os ortlogos retm habitualmente a mesma funo ao longo da
evoluo. A identificao de ortlogos critica na predio da funo genica em genomas
sequenciados.
- Gene parlogos genes relacionados, originados por duplicao dentro de um genoma. Os
ortlogos mantm a mesma funo ao longo da evoluo, enquanto os parlogos
desenvolvem novas funes (neofuncionalizao) ainda que relacionados ou prximos da
funo original ou parte das funes do gene original (subfuncionalizao). Levam a doenas
gnicas pois uma duplicao divergente pode alterar a estrutura que tinha inicialmente.
2. Material gentico, ciclo celular e ciclo biolgico_________________
Fases G1, S, G2 e M do ciclo celular
Fig. 6: Fases do ciclo celular
Sebenta de Gentica
FCML
13
Noo de ponto de restrio
Ponto de restrio: momento do ciclo celular em que as clulas ficam comprometidas para
entrar na Fase S. O ponto de restrio um checkpoint do ciclo celular fase G1 (R1). Clulas
que passam este ponto de restrio entram na fase S do ciclo celular. Os checkpoints do
ciclo celular so mecanismos de controlo que asseguram uma fidelidade da diviso celular
nas clulas eucariotas. Cada checkpoint verifica se os processos de cada fase do ciclo
celular foram completos de forma adequada antes de entrar na fase seguinte. O ponto de
restrio da fase G1 o primeiro checkpoint pelo qual as clulas passam no ciclo celular e
determina a passagem da clula para a fase S iniciando depois a diviso celular, o atraso na
entrada da fase S ou mesmo a passagem para um estado de interfase por tempo indefenido-
G0. Este ponto de restrio controlado essencialmente pela aco do CKI- p16 (p16 inibidor
da CDK). Esta protena inibe a CDK4/6 garantindo que esta no interage com a ciclina D1
que promove a progresso do cilco celular. A ciclina D induzida por factores oncognicos ou
factores de crescimento aumenta a sua expresso formando com CDK4/6 um complexo que
fosforila o retinoblastoma supressor de tumor que estimula o factor de transcrio E2F. Este
factor de transcrio aumenta a expresso da ciclina E que interage com CDK2 e promove a
passagem da fase G1 para a fase S.
G1/G0 quiescncia
Quando as clulas permanecem um longo tempo em G1, sem avanarem para a fase
subsequente do ciclo, designam-se por clulas em G0 ou quiescentes. Como exemplo da
variabilidade temporal de G1 refira-se que nas clulas do fgado pode ter uma extenso de
anos, em comparao com o que se verifica nas clulas da medula ssea em que a fase G1
tem uma durao de cerca de 20 horas e nas fases mais precoces das clulas embrionrias
em que a fase G1 est praticamente ausente. A deciso de uma clula permanecer em G0
ou entrar em diviso depende de factores extracelulares (ex. condies nutritivas) e celulares
(ex. tamanho celular suficiente para originar duas clulas). Na fase G0, as clulas diminuem
de tamanho, as proteinas e o RNA degradados no so rapidamente substitudos, a sntese
de macromolculas mais lenta, a actividade enzimtica e transporte transmembranar so
baixos e os ribossomas raramente se apresentam como polissomas.
A passagem de clulas de G0 para G1 permite que estas entrem no ciclo. Em clulas G0,
sujeitas a um estmulo extracelular, ocorre a activao de genes de activao precoce, cuja
transcrio se verifica alguns minutos aps a estimulao por factores de crescimento, sem
prvia sntese proteca. No grupo dos genes de activao precoce encontram-se os
protooncogenes FOS e JUN
Sebenta de Gentica
FCML
14
Controlo da proliferao celular e cascatas de transduo de sinal
O controlo preciso do ciclo celular exige uma complexa rede de vias de sinalizao que
integram sinais extra e intracelulares. No controlo da diviso celular intervm cinases
heterodimricas constitudas por ciclinas (sub-unidade reguladora) e cinases dependentes de
ciclinas (sub-unidade cataltica) = CDKs; e ainda protenas inibidoras das CDK (CDKIs). As
ciclinas activam as CDKs formando complexos moleculares Ciclinas- CDK. Estes
heterodmeros (MPFs-mitosis promoting factors) regulam a actividade de outras protenas
cruciais no ciclo celular, fosforilando-as nos seus locais reguladores promovendo a sua
activao ou desactivao. ainda importante mencionar os inibidores da CDK (CDKI) que
controlam a transio entre fases do ciclo celular, bloqueando a actividade das CDK; e o ciclo
de destruio de protenas (as j usadas e as que esto a inibir a progresso fase seguinte)
ciclo da Ubiquitina/Proteossoma.
Quadro. 1: tipos de ciclinas e CDK em cada fase do ciclo celular
Regulao das Ciclinas-CDK por CDKIs:
- p21CIP, p27 KIP2, p57 KIP2 inibem a actividade da Ciclina A-CDK2, e devem ser
degradadas antes de comear a replicao do DNA.
- INK 4S inibidores da cinase 4, inclui protenas que interagem com CDK4, CDK6, que ao
ligarem a INK4 bloqueiam a sua ligao Ciclina D. Controlam especificamente o princpio
de G1. ex. p16INK4a, p15INK4b, p18 INK4c e p19INK4d
Os sinais extracelulares podem ser mitognios, hormonas ou factores de crescimento e vo
promover a trancrio de dois tipos de genes:
Genes de resposta precoce:
- Induzidos por cascatas de transduo de sinal, que activam factores de transcrio pr-
existentes no citosol e no ncleo, Ex. c-fos, c-jun
- Esto presentes em G0
- Activados por fosforilao ou remoo de inibidor
Genes de resposta tardia:
- Genes de resposta precoce estimulam a transcrio de genes de resposta tardia
- No so transcritos em G0
Sebenta de Gentica
FCML
15
- Codificam factores de transcrio (ex. E2F), ciclinas tipo D e E, CDK2, CDK4, CDK6.
Regulao a nvel da fase G1:
O E2F- factor de transcrio codificado por genes de resposta tardia- activa genes que
codificam protenas envolvidas na sntese de DNA, ciclinas E e A e CDK2; autoestimula a
transcrio dos seus prprios genes e repressor da transcrio quando ligado protena
Rb, que liga complexos de Histona deacetilase. A Protena RB produto do gene supressor
tumoral RB est hipofosforilada no incio de G1 e o substracto mais importante dos
complexos ciclina-CDK em G1. A sua fosforilao previne a associao com E2F, no
impedindo a actividade deste ltimo. A fosforilao de Rb iniciada por ciclinaD-CDK4/6 e
terminada por ciclinaE-CDK2. Como consequncia: aumento da acumulao de ciclina E e
CDK2 (aco de E2F) e fosforilao de Rb mesmo na ausncia de ciclinaD-CDK4/6!
Fig. 7: Regulao Rb/E2F
Para que haja passagem pelo Ponto de Restrio necessrio que:
- os estmulos externos interajam com receptores desencadeando vias de transduo de
sinal, que chegam ao ncleo
- expresso dos complexos Ciclina-CDK de G1
- activao de Factores de Transcriio (ex. E2F) que promovem a transcrio de genes que
codificam protenas necessrias para sntese de DNA e que codificam as ciclinas e as CDKs
da Fase S (Ciclina A e E e CDK2)
- degradao dos inibidores da Fase S (CDKIs) por fosforilao e poliubiquitinao pela SCF
ubiquitina ligase e respectiva degradao no proteassoma (Ciclina A-CDK2)
Regulao de Rb e E2F na transio da fase Fase G1 para S:
E2F induz transcrio de ciclina A na transio entre G1 e S, formando complexos estveis,
impedindo a sua ligao ao DNA. As CDK2 so activadas pela fosfatase Cdc24A, no final de
G1. As ciclinas de G1 activam E2F via fosforilao de Rb e as ciclinas A--CDK2 inactivam-no
por fosforilao. A ciclina E-CDK2 tambm pode contribuir para a activao de complexos
pr-replicao: degradao da Ciclina E. A ciclina A--CDK2 marca a iniciao da sntese de
DNA e a traniio definitiva para S.
Sebenta de Gentica
FCML
16
Regulao a nvel da fase S:
Depois de haver a transio definitiva para a fase S d-se a sntese de ciclinas mitticas (final
de S) e a degradao, no proteossoma, dos inibidores da Fase S que liberta os complexos
ciclina-CDKs da Fase S (Ciclina ACDK2), activando-os. D-se a fosforilao de locais
reguladores de protenas que vo formar os complexos de pr-replicao de DNA dando
inicio replicao do DNA e produo de complexos ciclinaA-CDK2 mitticos fosforilados.
Regulao de Rb e E2F na passagem da fase G2 para M:
CDK1 a principal CDK em G2 e M e associa-se s ciclinas mitticas A e B. Esta CDK1
activada pela remoo do fosfato inactivador pela Cdc25C. a fosforilao de vrios
substractos que asseguram a transio G2/M.
Regulao da fase M:
Temos ento a activao, por desfosforilao, dos complexos Ciclina B-CDK1 mitticos e a
fosforilao de protenas envolvidas em processos de:
- Condensao cromosssmica
- Retraco do envelope nuclear
- Organizao do fuso acromtico
- Alinhamento dos cromossomas na placa metafsica
No final da mitose vamos ter os seguintes processos:
- O complexo promotor da anafase (APC): ubiquitina ligase, marca protenas reguladoras-
chave para degradao no proteassoma.
- Poliubiquitinao da securina d-se no incio da anafase, com a libertao dos cromatdeos-
irmos.
- Ubiquitinao das ciclinas B mitticas pelo APC
- Inactivao da actividade de cinase das CDK1 associadas s ciclinas mitticas
- Remoo de grupos fosfato adicionados a protenas por fosfatases-descondensao dos
cromossomas, reconstituio do envelope nuclear, reunio do complexo de Golgi
- Clula fica em G1 ou em G0.
Checkpoints:
A passagem nestas fases irreversvel, por serem marcadas pela degradao de protenas,
processo este tambm irreversvel.
Sebenta de Gentica
FCML
17
Fig. 8: Checkpoints
G1 check-point ou Ponto de Restrio:
Perodo do ciclo, no final de G1, onde a clula toma a deciso (ou no) de replicar o seu
DNA. Assegura que a clula possui a dimenso suficiente para se dividir, que existem
nutrientes em quantidade para suportar as clulas filhas resultantes e que no h danos nos
cromossomas. A deciso tomada mediante a presena de substncias indutoras
provenientes de outras clulas.
G2 check-point:
Assegura que a replicao correu bem e que o DNA no tem erros. As clulas que no
tenham replicado os seus cromossomas no entram na fase M. Envolve o reconhecimento de
DNA no-replicado e a inibio de MPF. Ex. ATR e Chk1 inibe a fosfatase Cdc25C logo h
inibio da ciclina A/B-CDK 1 e no se inicia a Fase M.
Check-point da Metafase:
Assegura que todos os cromossomas se encontram associados a um microtbulo
cinetocorial. Ex. Mad2 associa-se ao complexo cinetocorial que no est associado a
microtbulos, e inibe Cdc20, o que impede a poliubiquitinao da securina pela APC,
impedindo a anafase.
Fase S e replicao semi-conservativa e semi-descontnua do DNA
Replicao semi-conservativa do DNA:
um processo semi-conservativo.
Inicia-se em pontos especficos denominados unidades de replicao (originando o garfo
de replicao).
Sempre na direco 5 3.
um processo semi-descontnuo.
Participam vrias classes de protenas
Sebenta de Gentica
FCML
18
Quadro 2: Tipos de DNA polimerase
PCR:
um mtodo para a amplificao de sequncias-alvo de DNA definidas, presentes numa
preparao de DNA. utilizado DNA genmico ou uma populao completa de cDNA.
necessria informao prvia a respeito das sequncias-alvo para permitir desenhar dois
oligonucletidos iniciadores- primers que limitam a zona a ser ampliada. Estes primers vo
ligar-se cadeia complementar sua sequncia.
Fig. PCR
Consiste numa srie de ciclos de trs reaces sucessivas:
- Desnaturao- que ocorre entre 93-95 no DNA humano (cromossomas em interfase)
- Emparelhamento- entre 50-70.
Sebenta de Gentica
FCML
19
- DNA sntese- que ocorre entre 70-75, e utiliza uma DNA polimerase termoestvel (estvel
ao calor) como por exemplo a Taq DNA polimerase.
Limitaes:
- Necessidade de conhecimento prvio da sequncia-alvo.
- A dimenso dos produtos amplificados- pequena
- A quantidade limitada de material que pode ser clonado.
- Infidelidade na replicao do DNA.
- Contaminao por DNA estranho
Vantagens:
- Determina sequncias de DNA a partir de quantidades muito reduzidas de DNA
- um mtodo rpido e fcil de executar.
- um mtodo sensvel.
- um mtodo fivel mesmo em tecidos ou clulas degradadas.
Aplicaes:
- Deteco de polimorfismos, mutaes pontuais, infeces virais e bacterianas
- Medicina Forense.
- Linkage Gentico.
Sequenciao do DNA:
Os fragmentos de DNA usados na sequenciao podem ser obtidos tanto por clonagem
como por PCR. O DNA pode ser sequenciado por mtodos quimcos (Mtodo de Maxam and
Gilbert) mas a sequenciao do DNA por mtodo enzimtico (Mtodo de dideoxy),
actualmente o mais utilizado. Os dois mtodos exigem electroforese com capacidade para
diferenciar fragmentos de DNA em que o tamanho diverge apenas por uma base.
Mtodo Dideoxy: o DNA fornecido em cadeia simples, e actua como molde para a sntese
duma cadeia complementar pela DNA polimerase. Efectuam-se 4 reaces paralelas, cada
uma contendo os 4 dNTP, mais uma pequena proporo de um dos 4 didesoxinucletidos
(ddNTPs) anlogo, que actua como finalizador da cadeia. Marca-se um dos 4 dNTPs ou o
primer com um radioistopo ou um fluorocromo, de modo que o DNA sintetisado fique
marcado. Os fragmentos so separados numa electroforese com gel desnaturante de
polyacrylamida.
Sequenciao automtica: utiliza marcadores fluorocromos diferentes para cada uma das 4
reaces, marcando os ddNTPs ou o primer. As 4 reaces podem ser efectuadas
juntas,devido colorao diferencial dos ddNTPs. Durante a electroforese um monitor
Sebenta de Gentica
FCML
20
detecta e recorda o padro de fluorescncia diferente para cada um dos fluorocromos, e
armazena esta informao num computador.
Aplicaes da sequenciao:
- deteco de mutaes.
- deteco de polimorfismos (genotipagem).
- identificao de novos genes e a elucidao da sua sequncia.
Normal Mutao (185del A)
Protenas estabilizadoras da cadeia simples, helicases e topoisomerases 1 e 2
Helicase: uma enzima utilizada para separar as duas cadeias de DNA que formam a dupla
hlice utilizando energia da hidrlise do ATP e actuando ao nvel da quebra das pontes de
hidrognio entre as bases azotadas. H 24 tipos de helicases nos seres humanos uma vez
que h vrias situaes em que necessrio separar as cadeias da dupla hlice.
Topoisomerase: ajuda no desenrolamento da dupla hlice de DNA atravs da quebra das
ligaes de fosfato. A topoisomerase I actua sobre uma cadeia de DNA enaquanto que a
topoisomerase II actua em ambas as cadeias de DNA. As topoisomerases tambm esto
envolvidas em mecanismos de reparao de DNA.
O problema do fim da replicao: telomerase
Sequencing of telomeres from a dozen or so organisms, including humans, has shown that
most are repetitive oligomers with a high G content in the strand with its 3 end at the end of
the chromosome. The telomere repeat sequence in humans and other vertebrates is
TTAGGG. These simple sequences are repeated at the very termini of chromosomes for a
total of a few hundred base pairs in yeasts and protozoans and a few thousand base pairs in
vertebrates. The 3 end of the G-rich strand extends 1216 nucleotides beyond the 5 end of
the complementary C-rich strand. This region is bound by specific proteins that both protect
start
Sebenta de Gentica
FCML
21
the ends of linear chromosomes from attack by exonucleases and associate telomeres in
specific domains within the nucleus. The need for a specialized region at the ends of
eukaryotic chromosomes is apparent when we consider that all known DNA polymerases
elongate DNA chains at the 3 end, and all require an RNA or DNA primer. As the growing fork
approaches the end of a linear chromosome, synthesis of the leading strand continues to the
end of the DNA template strand, completing one daughter DNA double helix. However,
because the lagging-strand template is copied in a discontinuous fashion, it cannot be
replicated in its entirety. When the final RNA primer is removed, there is no upstream strand
onto which DNA polymerase can build to fill the resulting gap. Without some special
mechanism, the daughter DNA strand resulting from lagging-strand synthesis would be
shortened at each cell division. The problem of telomere shortening is solved by an enzyme
that adds telomeric sequences to the ends of each chromosome. The enzyme is a protein and
RNA complex called telomere terminal transferase, or telomerase. Because the sequence of
the telomerase-associated RNA serves as the template for addition of deoxyribonucleotides to
the ends of telomeres, the source of the enzyme and not the source of the telomeric DNA
primer determines the sequence added. Telomerase is a specialized form of a reverse
transcriptase that carries its own internal RNA template to direct DNA elongating the 3 end of
the single-stranded DNA at the end of the G-rich strand. The absence of telomere DNA results
in adverse effects, including fusion of chromosome termini and chromosomal loss.
Para alm da telomerase, existe outro processo atravs do qual os tumores conseguem
fazer de forma alternativa a manuteno dos telmeros: o ALT.
No ALT, uma cadeia de DNA de um telmero liga-se a uma cadeia complementar de outro
telmero, obtendo assim o modelo para a sntese do novo DNA telomrico. Abrem-se as
duas cadeias como se v no esquema seguinte, servindo uma de molde outra. Aps a crise
celular, alguns telmeros ficam to curtos que as repeties telomricas encontram-se logo
aps a zona subtelomrica do cromossoma, onde j no existem as repeties da sequncia
telomrica. Quando se d o mecanismo ALT, possveis sequncias GGTTAG perdidas nas
zonas subtelomricas podem agir como primer de adio de novo DNA telomrico,
resultando isto na substituio das variaes subtelomricas por repeties telomricas
GGTTAG. Os telmeros mantidos pelo ALT apresentam elevada heterogeneidade e so
observados em clulas que tambm apresentam telomerase.
Fase M
Condensao de cromatina- ver regulao e organizao da cromatina (pg. 41)
Sebenta de Gentica
FCML
22
Mitose
Fuso acromtico e protenas contrcteis do citoplasma
Designa-se por fuso acromtico ou mittico a estrutura dinmica que se forma durante a
mitose, fundamentalmente constituda pelos microtbulos e centrosomas. A formao do fuso
acromtico inicia-se na prometafase, completa-se na metafase e dissipa-se, graudalmente,
na anafase B. O fuso morfologicamente constitudo por dois plos diametralmente opostos,
sendo cada um formado por uma regio polar onde se localiza um centrossoma formado por
dois centrolos que se encontram situados a curta distncia um do outro em posio
ortogonal. Os polos esto interligados entre si por um conjunto de microtubulos, muitos dos
quais esto directamente ligados ao cinetocoro dos cromossomas. Entre as duas regies
polares, numa posio perpendiclar aos microtbulos e, sensivelmente a igual distncia entre
os dois polos, encontra-se uma zona complanar que, genericamente, se chama plano
equatorial do fuso (tambm conhecida por placa equatorial ou mittica) o qual separa o fuso
acromtico em duas zonas iguais. Associado aos microtbulos encontra-se um grupo de
Sebenta de Gentica
FCML
23
protenas que participam em muitos processos celulares e que tm um papel central na
formao e funo do aparelho mittico. Estas protenas, denominadas protenas motoras,
pertencem a duas famlias: as cinesinas e as dineinas. Estas protenas tm varios domnios,
nomeadamente um domnio para a formao de dmeros que permita a ligao aos
microtbulos, e um outro domnio para a associao com outros componentes celulares.
Estas protenas ligam os microtbulos e utilizam a energia proveniente da hidrlise do ATP
para mover estruturas celulares tanto na direco (-) no caso das dineinas, como na direco
(+) no caso das cinesinas. No incio da fase S, junto a cada centrolo, forma-se um esboo
microtubular chamado procentrolo. Os esboos microtubulares de cada procentrolo
organizam-se e crescem por adio de monmeros de tubulinas. Os tripletos microtubulares
atingem a sua organizao centriolar completa geralmente no fim da fase S. Cada centrolo
fica acompanhado de um novo centrolo ficando a existir 2 centrossomas (4 centrolos) na
mesma regio cromossmica. Quando se inicia a mitose, cada centrossoma comea a
deslocar-se para uma posio diametralmente oposta, atingindo essa posio na metafase
quando se completa o fuso acromtico.
Recombinao mittica e troca de cromtides irms (SCEs)
Recombinao mittica: most spontaneous RCOs (reciprocal crossovers) are initiated by DNA
double-strand breaks (DSBs). Recombinational repair of a DSB requires a template; when the
homologous chromosome serves that role, it provides the opportunity for an RCO. Since there
is also evidence that single-stranded nicks and gaps are recombinogenic, it is likely that
several types of DNA lesions may be important for spontaneous mitotic recombination events.
In addition, some recombinogenic agents (such as ultraviolet radiation) are thought to produce
nicks that result in DSBs when the nicked DNA is replicated. Thus, the question of why
becomes tied up with the question of when.Whereas meiotic recombination occurs during
meiosis, most mitotic recombination probably does not occur during mitosis, but during
interphase.
A SCE (Sister Cromatid Change) consiste numa troca simtrica e exacta de material gentico
entre cromtides irms. Estas trocas ocorrem em qualquer mitose, sendo mais frequentes
contudo na presena de agentes lesivos do genoma. Pensa-se que podem ter um papel de
reparao. Este fenmeno no acarreta nenhuma alterao morfolgica ou funcional no
cromossoma, desde que no se trate de uma troca desigual, j que, como foi dito, envolve
uma troca entre pores idnticas de DNA. A frequncia de SCE muito elevada na
sndroma de Bloom.
O ciclo biolgico e a reduo meitica da ploidia
Sebenta de Gentica
FCML
24
Meiose
Compreende duas divises celulares sucessivas mas somente um ciclo de replicao de
DNA de modo a qua as clulas filhas finais so haplides. A primeira diviso (meiose I)
uma diviso de reduo (heterotpica) produzindo duas clulas haplides oriundas de uma
nica clula diplide. A segunda diviso (meiose II) uma diviso equacional (homotpica)
que separa os cromatdios irmos das clulas haplides. A meiose faz parte de um ciclo
celular designado ciclo celular meitico que corresponde a duas fases distintas: interfase pr-
meitica e a fase M (meiose). Ver pgina 397 do Carlos Azevedo.
Quiasmata e crossing-over
Durante a profase da meiose I, os cromossomas sinpticos em cada bivalente (cromossoma
materno unido ao cromossoma homlogo paterno), trocam segmentos entre si de forma
aleatria. No estgio de zigoteno (ver meiose), cada par de homlogos comea a formar um
complexo sinaptonmico que consiste em dois cromossomas intimamente emparelhados,
separados por um longo eixo linear de protenas. A finalizao desse complexo marca o incio
do estgio de paquiteno, que quando ocorre a recombinao- crossing-over). O crossing-
over envolve a quebra fsica da dupla hlice de uma cromtide paterna e uma materna e a
troca de informao gentica entre elas. A recombinao resulta ento da troca de grupos de
genes produzida por ruptura e nova fuso de fragmentos anlogos correspondentes a
cromatdeos internos no irmos de um bivalente. Em conjunto, a recombinao entre
homlogos na profase I e a distribuio independente dos homlogos na anafase I assegura
que cada indivduo possa produzir um nmero quase ilimitado de gmetas geneticamente
diferentes. O mecanismo que proporciona o alinhamento dos homlogos no est
determinado. Entretanto, considera-se que essa aposio ntima seja necessria para a
recombinao. Supe-se que os ndulos de recombinao- grandes acumulados de mltiplas
protenas localizados a intervalos especficos no complexo sinaptonmico- realizem a
mediao nos eventos da recombinao. Pode-se ver que os dois homlogos esto
fisicamente em contacto em pontos especficos. Cada conexo considerada um quiasma
(ou quiasmata) e marca um ponto de crossing-over. Alm do seu papel na recombinao,
supe-se que os quiasmas sejam essenciais para a segregao cromossmica correcta na
meiose I. Ao manter os homlogos materno e paterno de cada par de cromossomas juntos no
fuso at anafase I, eles tm um papel anlogo ao dos centrmeros na mitose e meiose II.
H evidncias genticas de que crianas com um nmero errado de cromossomas
frequentemente resultam de gmetas provenientes de uma clula na qual um bivalente no
sofreu crossing-over.
Sebenta de Gentica
FCML
25
Converso e complementao gnica
Converso gnica: fenmeno associado existncia de sequncias homlogas. Na
converso gnica, uma sequncia modificada a partir de outra, provavelmente por
formao de heteroduplexes entre as duas cadeias de DNA. O mecanismo de reparao de
erros de emparelhamento (MMR) reconhece uma das cadeias como errada e procede
correco a partir da outra cadeia.
Fig. 9: Converso gnica
Complementao gnica: refere-se hibridao entre 2 genomas com dfice em
determinados genes diferentes, sendo o resultado obtido um genoma normal por
complementariedade. Dois exemplos so a doena de Hurler e a doena de Hunter, em que
o fentipo pode ser normal se os defeitos no forem allicos (ver pg. 110).
3. Genes e expresso do material gentico_______________________
Sebenta de Gentica
FCML
26
Transcrio
RNA polimerases I, II e III
A RNA polimerase constituda por 4 subunidades 2o, |, | e o que compem a
holoenzima. A subunidade o responsvel pela identificao de um local promotor local de
reconhecimento onde a transcrio tem incio, actuando apenas durante a iniciao da
transcrio, constantemente reciclada, existindo em muito menores concentraes nas
clulas, que a enzima mnima. Existem diversas subunidades o que reconhecem diferentes
sequncias de consenso, caractersticas de genes diferentes. A RNA polimerase sem a
subunidade o designada por core enzima (2o, |, |). Estas subunidades constituem o local
cataltico, sendo a subunidade | responsvel pela ligao cadeia de DNA padro e a |
pela ligao aos substratos ribonucletidos trifosfatados. Alm destas subunidades convm
tambm referir o factor rho (Erro! Marcador no definido.), responsvel pela terminao.H
trs tipos de RNA polimerase descritas no quadro seguinte:
Fig. 10: Expresso do material gentico numa clula animal
Sebenta de Gentica
FCML
27
Quadro 2: Trs tipos de RNA polimerase dos eucariticos
Locais de iniciao
As RNA polimerases dos eucariotas no tm autonomia para iniciar a transcrio. preciso
que combinaes de elementos, com sequncias curtas, adjacentes ao gene, actuem como
sinais de reconhecimento para que os factores de transcrio possam ligar-se ao DNA para
guiar e activar a polimerase. Frequentemente, um conjunto desses elementos com
sequncias curtas fica agrupado a montante da sequncia codificadora de um gene
constituindo colectivamente, o promotor. Depois de vrios factores comuns de transcrio se
ligarem regio do promotor, uma RNA-polimerase liga-se ao complexo desses factores e
activada para iniciar a sntese de RNA a partir de um nico ponto.
Elementos cis e elementos trans
Diz-se que os factores de transcrio so trans-actuantes pois so sintetizados por genes
situados em locais distantes, migrando para os seus locais de aco. Os elementos
promotores, reforadores e silenciadores por sua vez so cis-actuantes uma vez que a sua
funo se limita ao local de DNA onde se encontram.
Locais de ligao TATA, GC, CAAT e factores de transcrio
Os elementos promotores dos genes que so transcritos activamente pela RNA polimerase II,
incluem sempre uma TATA box, frequentemente a TATA-AA ou uma variante, situada cerca
de 25pb a montante (-25) do local de incio da transcrio. Uma mutao no elemento TATA
no impede o incio da transcrio, mas desloca o seu ponto de incio da sua posio normal.
Os promotores de muitos outros genes, inclusive os dos genes de manuteno, no tm
TATA boxes, mas, frequentemente tm uma GC box contendo varitantes da sequncia
consensual GGGCGG. Outro elemento promotor frequente a CAAT box, situado
aproximadamente a -80 que usualmente o maior determinante da eficincia do promotor.
Note porm que, embora as sequncias da GC box e da CAAT box sejam assimtricas,
Sebenta de Gentica
FCML
28
ambas parecem capazes de funcionar em qualquer orientao. Alm dos elementos comuns
de transcrio, localizados a montante e reconhecidos por factores de transcrio ubquos,
existem elementos de reconhecimento mais especficos, que s so identificados por factores
de transcrio restritos a determinados tecidos. (ver pgina 42)
Fig. 11: Promotores eucariticos. chevron orientation: > = normal orientation; < = reverse orientation.
Maturao do RNA:
Faz parte da regulao ps-transcripcional.
F Fo or rm ma a o o d do o c ca ap p
Ocorre logo aps a transcrio. No caso dos transcritos primrios, que sero processados
para produzir mRNA, um nucleosdeo metilado, a 7-metilguanosina (m
7
G), ligado ao
primeiro nucletido 5 do transcrito de RNA por uma ligao fosfodister especial 5-5. Tendo
em conta que o carbono 5 do resduo m
7
G e o carbono 5 do primeiro nucletido so
efectivamente ligados, diz-se que a extremidade 5 est bloqueada ou capeada (cap). Os
transcritos dos genes de snRNA tambm so capeados, mas as suas capas podem sofrer
ainda outras modificaes. Supe-se que o cap possa ter as seguintes funes:
- proteger o transcrito do ataque da exonuclease 5 3 (molculas de mRNA sem cap
so desnaturadas rapidamente)
- facilitar o transporte do ncleo para o citoplasma
- facilitar o encadeamento de RNA
- desempenhar um papel importante no encaixe da subunidade menor (40S) dos
ribossomas citoplasmticos no mRNA
Sebenta de Gentica
FCML
29
P Po ol li i- -a ad de en ni il la a o o
A transcrio pelas RNA-polimerases I ou II pra logo que a enzima reconhece um stio
especfico de fim da transcrio. No entanto, este reconhecimento dificultado porque as
extremidades 3 das molculas do mRNA so determinadas por uma reaco de clivagem
posterior transcrio. A sequncia AAUAAA o elemento principal para assinalar a
clivagem 3 na grande maioria dos transcritos por polimerase II (os transcritos dos genes de
histonas e dos genes de snRNA so excepes importantes). A clivagem ocorre num stio
especfico, localizado a jusante do elemento AAUAAA. Em busca do ponto de clivagem, a
transcrio pode continuar por muitos nucletidos at que termine alguns stios depois. Nas
clulas de mamferos, uma vez ocorrida a clivagem a jusante do elemento AAUAAA, cerca de
200 resduos de adenilato (ex. AMP) so adicionados sequencialmente pela enzima poli(A)-
polimerase para formar uma cauda poli(A). Supe-se que essa cauda tenha vrias funes:
- facilitar o transporte das molculas de mRNA para o citoplasma
- estabilizar ao menos parte das molculas de mRNA no citoplasma (o encurtamento da
extenso da poli(A) est associado com a degradao do mRNA, mas algumas espcies
de mRNA (ex. o mRNA da actina) permanecem estveis com pouca ou nenhuma poli(A)
- facilitar a traduo, ao permitir uma intensificao do reconhecimento do mRNA pela
maquinaria ribossmica
I In nt tr r e es s e e e ex x e es s
As sequncias codificadoras da maioria dos genes dos vertebrados, tanto os que codificam
polipptidos quanto os que codificam outras molculas de RNA que no o mRNA, so
divididas em segmentos- os exes- que so separados por sequncias intercaladas no
codificadoras- os intres. A transcrio consiste na produo de uma sequncia de RNA
complementar ao comprimento total do gene, abrangendo exes e intres. Frequentemente o
RNA transcrito sofre splicing.
s sn nR RN NA As s, , s sp pl li ic ce eo os ss so om ma a e e s sp pl li ic ci in ng g d do os s R RN NA As s. . S Sp pl li ic ci in ng g a al lt te er rn na at ti iv vo o
Splicing: Relativamente ao splicing, este processo consiste na introduo de alteraes na
sequncia original de exes. Desta forma, a clula pode gerar diferentes mRNA a partir de
um nico gene e consequentemente produzir diferentes protenas. O mecanismo de splicing
de RNA dependente da identidade das sequncias de nucletidos nos limites exes/intres
(junes de splicing). particularmente sensvel ao que tem sido denominada a regra GT-
AC: os intres quase sempre comeam com GT (GU ao nvel do RNA) e terminam com AG.
Embora os dinucletidos conservadores GU e AG sejam cruciais no splicing, no so
suficientes para indicar a presena de um intro. As sequncias adjacentes aos dinucletidos
Sebenta de Gentica
FCML
30
GT e AC mostram um grau considervel de conservao. Existe uma terceira sequncia
conservada nos intres que funcionalmente importante para o splicing, o local de
ramificao, quase sempre localizado perto do final do intro, antes do dinucletido terminal
AG. O mecanismo de splicing o seguinte:
- clivagem na juno da cadeia 5
- ataque nucleoltico ao A invariante do stio de ramificao pelo nucletido G terminal do stio
dador de splicing, a fim de formar uma estrutura em forma de lao
- clivagem na juno da cadeia 3, com a libertao do RNA do intro sob a forma de um lao
e o splicing dos exes do RNA.
Estas reaces so mediadas por um complexo de RNA e de protenas, o spliceosoma, que
consiste em cinco tipos de snRNA (small nuclear RNA) e de mais de 50 protenas.
Cada molcula de snRNA est ligada a protenas especficas, formando partculas de snRNA
(small nuclear ribonucleoproteins) e a especificidade da reaco de splicing estabelecida
pela formao de pares de bases RNA-RNA entre o RNA transcrito e as molculas de
snRNA. Na grande maioria dos intres de genes que codificam polipptidos, as cinco
espcies de snRNA so U1, U2, U4, U5, U6. O terminal 5 do snRNA U1 tem uma sequncia
UACUUAC, cujas bases emparelham com a sequncia de consenso do stio dador de
splicing (GUAAGUA). Depois da snRNP U1 se ligar, o snRNA U2 reconhece o local de
ramificao por uma reaco semelhante ao emparelhamento de base e logo aps, a
interaco entre os snRNP U1 e U2 aproxima intimamente as duas extremidades que sero
unidas. Em seguida, uma partcula com mltiplos snRNP, contento snRNAs U4 U5 e U6
associa-se ao complexo snRNPs U1-U2. Supe-se que o spliceosoma age de modo
progressivo e depois de reconhecer um local de splicing 5 percorre a sequncia de RNA at
encontrar o local de splicing 3.
Fig. 12: (A) Mecanismo de splicing (B) Papel dos snRNPs.
Sebenta de Gentica
FCML
31
Stios cripticos de splicing: Sequncias que so muito semelhantes s sequncias
consensuais dos stios dador ou receptor de splicing podem, coincidentemente existir em
intres e exes. Essas sequncias no so normalmente utilizadas no splicing e, por isso,
so conhecidas como stios crpticos de splicing. Uma mutao pode activar um destes stios
ao tornar a sequncias mais parecida com a sequncia consensual do stio dador ou receptor
de splicing e o stio crptico pode, ento, ser reconhecido e utilizado pelo spliceosoma. Um
exemplo pode ser visto na figura seguinte que mostra uma mutao identificada num
paciente com distrofia muscular dos membros. A mutao foi encontrada no gene da calpaina
3, um lcus conhecido para esse tipo de distrofia muscular, mas ocorreu na posio da
terceira base de um codo e parecia ser uma mutao silenciosa (subsitutio de um codo
de glicina por outro de glicina tambm). No entanto, acredita-se que, a mutao seja
patognica uma vez que a substituio resulta na activao de uma sequncia dadora de
splicing, oculta no exo 16, resultando num splicing aberrante, com a perda da sequncia
codificadora do exo 16 e a introduo de uma mudana na fase de leitura.
Fig. 13: Exemplo de stio crptico de splicing
Splicing alternativo: A large percentage of human genes undergo alternative splicing whereby
different exon combinations are included in transcripts from the same gene during RNA
processing. For many genes, numerous isoforms can be generated at the RNA level, but often
the functional significance is poorly appreciated. In some genes, alternative splicing results in
very considerable diversity in the untranslated regions. For example, in the liver alternative
splicing results in at least 8 different 5 UTR sequences for human growth hormone receptor
mRNA, but the functional significance, if any, is not understood. Alternative splicing of coding
sequence exons is also common and some of the resulting protein isoforms have been shown
to be tissue specific, so that individual exons present in one isoform but not in others may be
termed muscle-specific , brain-specific etc. The different isoforms can provide a variety of
possibilities for altered functional properties but detailed knowledge of the functional
significance of the different isoforms is still comparatively sparse. The best understood model
system for understanding the regulation of splicing is the sex determination pathway in
Sebenta de Gentica
FCML
32
Drosophila which also controls gene dosage. Alternative splicing is used in each branch of this
pathway to control the expression of transcriptional regulators or chromatin-associated
proteins that influence transcription, and both positive and negative control of splicing is
evident. In mammalian cells candidate splice regulators are the SR family of RNA-binding
proteins (which have a distinctive C terminal domain rich in serine (S)-arginine (R) dipeptides]
and some HnRNP (heterogeneous nuclear ribonucleoprotein particle) proteins. These proteins
are known to promote various steps in assembly of spliceosomes and they are also known to
bind to splicing enhancer sequences, regulatory sequences which can enhance splice site
recognition. Aumenta a capacidade para originar diversidade proteica.
E Ed di i o o d do o R RN NA A: :
uma forma de processamento posterior transcrio que pode envolver a insero ou
deleco de nucletidos, mediada por enzimas, ou a substituio de nucletidos individuais a
nvel de RNA. A expresso do gene da apolipoprotena B humana no intestino envolve a
edio do RNA tecido-especfica (figura seguinte). O codo 2153, especificado pelo tripleto
CAA na posio 6666-6668 especfico para a glutamina na ApoB100 sintetizada no fgado.
No intestino, porm, o codo CAA convertido, pela edio de RNA, no codo de terminao
UAA, resultando num produto mais curto- ApoB48.
Fig. 14: Exemplo de edio de RNA
RNAs no codificantes (ncRNA):
The term non-coding RNA (ncRNA) is commonly employed for RNA that does not encode a
protein, but this does not mean that such RNAs do not contain information nor have function.
Although it has been generally assumed that most genetic information is transacted by
proteins, recent evidence suggests that the majority of the genomes of mammals and other
complex organisms is in fact transcribed into ncRNAs, many of which are alternatively spliced
and/or processed into smaller products. These ncRNAs include microRNAs and snoRNAs
(many if not most of which remain to be identified), as well as likely other classes of yet-to-be-
Sebenta de Gentica
FCML
33
discovered small regulatory RNAs, and tens of thousands of longer transcripts (including
complex patterns of interlacing and overlapping sense and antisense transcripts), most of
whose functions are unknown. These RNAs (including those derived from introns) appear to
comprise a hidden layer of internal signals that control various levels of gene expression in
physiology and development, including chromatin architecture/epigenetic memory,
transcription, RNA splicing, editing, translation and turnover. RNA regulatory networks may
determine most of our complex characteristics, play a significant role in disease and constitute
an unexplored world of genetic variation both within and between species.
Fig. 15: Classes de ncRNA
microRNA e o mecanismo de interferncia de RNA
MicroRNA (miRNA) is a short (about 21 to 23 nucleotides) single-stranded RNA molecule that
is now recognized as playing an important role in gene regulation even though the term has
been in use only since 2001. It is similar to, but distinct from, another type of short RNA,
known as small interfering RNA (siRNA).
Mecanismo de interferncia de RNA: Long double-stranded RNAs (dsRNAs; typically >200 nt)
can be used to silence the expression of target genes in a variety of organisms and cell types
(e.g., worms, fruit flies, and plants). Upon introduction, the long dsRNAs enter a cellular
pathway that is commonly referred to as the RNA interference (RNAi) pathway. First, the
dsRNAs get processed into 20-25 nucleotides small interfering RNAs- siRNAs by an RNase
III-like enzyme called Dicer (initiation step). Then, the siRNAs assemble into
endoribonuclease-containing complexes known as RNA-induced silencing complexes
(RISCs), unwinding in the process. The siRNA strands subsequently guide the RISCs to
complementary RNA molecules, where they cleave and destroy the cognate RNA (effecter
step). Cleavage of cognate RNA takes place near the middle of the region bound by the
Sebenta de Gentica
FCML
34
siRNA strand. In mammalian cells, introduction of long dsRNA (>30 nt) initiates a potent
antiviral response, exemplified by nonspecific inhibition of protein synthesis and RNA
degradation. The mammalian antiviral response can be bypassed, however, by the
introduction or expression of siRNAs.
Fig. 16: Mecanismo de interferncia de RNA
Rede de regulao mediada pelos ncRNAs
ncRNA so os RNA no codificantes dos quais se destaca o miRNA (micro RNA) que um
RNA de regulao codificado por regies intrnicas e o siRNA. O mecanismo de
transferncia atravs do siRNA foi descrito anteriormente. O mecanismo de regulao
atravs do miRNA est descrito na figura seguinte:
Sebenta de Gentica
FCML
35
Mecanismo de regulao atravs do miRNA (fig. 21):
1) microRNAs so transcritos como parte de uma molcula de RNA: Pri-miRNA.
2) O pri-miRNA processados no ncleo pela ribonuclease Drosha Pre-miRNA.
3) Pre- miRNA transportado para o citoplasma pelo mecanismo exportin 5-dependent
4) No citoplasma, o Pre-miRNA digerido por uma ribonuclease- Dicer originando uma
cadeia de miRNA especfica e maturada.
5) O miRNA de cadeia simples vai ligar-se ao RNA- induced Silencing Complex (RISC) que
participa no RNAi (interferncia)
6) O miRNA+RISC vai unir-se ao mRNA (3UTR) impedidno a sua traduo resultando na
diminuio da expresso gnica do respectivo gene.
Aproximadamente 60% dos miRNAs so expressos independentemente, 15% so expressos
em clusters, e 25% em intres. Although miRNA and siRNA both have gene regulation
functions, there are subtle differences. miRNA may be slightly shorter than siRNA (which has
20 to 25 nucleotides). miRNA is single-stranded, while siRNA is formed from two
Sebenta de Gentica
FCML
36
complementary strands. The two kinds of RNA are encoded slightly differently in the genome.
And the mechanism by which they regulate genes is slightly different. miRNA attaches to a
piece of messenger RNA (mRNA) which is the master template for building a protein in a
non-coding part at one end of the molecule. This acts as a signal to prevent translation of the
mRNA into a protein. SiRNA, on the other hand, attaches to a coding region of mRNA, and so
it physically blocks translation. Os miRNAs so molculas endgenas de regulao enquanto
os siRNAs so molculas exgenas processadas atravs de RNA de cadeia dupla. No
entanto, ambos so processados por Dicer. A Dicer encaminha o produto para o RISC que
o efector do silenciamento/interferncia. O RISC pode silenciar mRNAs por 2 mecanismos:
1. Se h emparelhamento perfeito entre ncRNA (miRNA essencialemnte) e mRNA este
degradado por Argonauta (subunidade que degrada mRNA emparelhado com miRNA);
2. Se h erros de emparelhamento (mismatches) o mRNA pode ser localizado em focos
citoplasmticos de processamento (P-bodies) onde degradado ou sequestrado da
traduo.
Cdigo gentico
Degenerescncia do cdigo gentico
A sequncia de aminocidos de uma protena determinada pela sequncia de bases do
DNA correspondente. Esta correspondncia conseguida pela utilizao de grupos de trs
bases, denominadas codes, que correspondem a um nico aminocido. Deste modo, pela
utilizao de 4 bases combinadas, possvel obter 43 arranjos diferentes, ou seja, 64
diferentes codes. Se a cada codo corresponder um aminocido diferente e considerando
os 20 aminocidos normalmente utilizados na sntese proteica, observaramos que 44 das
combinaes no codificariam qualquer aminocido. No entanto, muitos dos aminocidos
podem ser codificados por mais de uma combinao, ou seja, o cdigo degenerado (com
excepo da metionina e da triptofana, todos ou outros aminocidos so codificados por
vrios tripletos sinnimos).
A quase universalidade do cdigo gentico
A universalidade do cdigo gentico tem como base o facto de que mRNA de clulas
distintas pode ser traduzido em sistemas heterlogos. No entanto, presentemente conhecem-
se limites universalidade; existem preferncias na utilizao de codes consoante o tipo
celular. Esta diversidade aplica-se igualmente aos tripletos que no codificam para nenhum
aminocido e que correspondem a sinais de paragem da traduo do mRNA. Uma excepo
universalidade o facto de nas mitocndrias existirem 3 codes que designam informao
Sebenta de Gentica
FCML
37
diferente relativamente ao citoplasma da mesma clula codo UGA um codo de
terminao no cdigo usual e codifica triptofana no cdigo mitocondrial; o codo AUA codifica
para a isoleucina no cdigo usual e para a metionina na mitocndria; o codo CUA codifica
para a leucina no cdigo usual e para a treonina na mitocndria.
Cdigo gentico mitocondrial
Diferente do DNA nuclear que forma longas fitas, constitudas cada uma por dupla hlice e
que codificam aproximadamente 100.000 genes, o mtDNA representa 1 a 2% do DNA celular,
em duplo filamento circular, codificando 37 genes. O mtDNA codifica aproximadamente 10%
das protenas constitutivas das mitocndrias, como consequncia, para um bom
funcionamento destas, necessria uma boa cooperao entre o DNA nuclear e o DNA
mitocondrial. O mtDNA no tem nada de especial na sua composio qumica que o
diferencie do DNA nuclear, entretanto, possui um cdigo gentico prprio. Por outro lado, a
sua organizao bastante econmica, j que apenas 10% da sua totalidade no
codificante. O seu genoma haplide devido sua herana ser estritamente materna, por
isso no est submetido a processos de recombinao. O mecanismo pelo qual a
mitocndria paterna excluda do embrio logo aps a fertilizao ainda no est bem
esclarecida. Os filamentos de DNA mitocondrial tm uma distribuio assimtrica de
guaninas e citosinas, o que gera uma cadeia pesada (H) e outra leve (L). Cada filamento
transcrito a partir de um promotor PL e PH1, localizados na regio controle, na qual se inclui
o D-Loop que uma regio gerada pela sntese de um segmento curto da cadeia pesada (H)
denominado 7SDNA, onde se encontra a origem da replicao da cadeia H. Esta regio
muito importante do ponto de vista forense, devido ao facto de ser uma regio hipervarivel e
suficientemente pequena para ser abordada por meio de seqenciamento por PCR.
Caractersticas do mtDNA:
- muito sensvel ao dano oxidativo causado principalmente por um grande nmero de
radicais livres proporcionado um ambiente favorvel a mutao do DNA.
- no possui histonas, que exercem um papel protector no DNA nuclear
- a mtDNA polimerase tm uma pobre actividade reparadora comparada com a nuclear.
- a reparao do DNA dependente de exciso de nucleotdeos no est presente
- a tpica estrutura D-Loop, onde h formao momentnea de fitas simples pode influenciar o
padro de mutao pontual, j que a taxa de despurinao do DNA de cadeia simples 4x
maior do que a do DNA cadeia dupla.
- codifica componentes do sistema da fosforilao oxidativa (complexo I, III, IV e V);
componentes de tRNA e rRNA.
Sebenta de Gentica
FCML
38
Todos estes fatores fazem com que o DNA mitocondrial tenha uma taxa de evoluo 5 a 10
vezes maior que o DNA nuclear, o que implica uma hipervariabilidade entre a populao
humana. A alta taxa de substituio de bases possibilita que seja observada uma ou mais
diferenas na sequncia nucleotdica das regies hipervariveis do mtDNA, quando
indivduos da linhagem materna de uma mesma famlia so comparados. Por outro lado, o
mtDNA est presente em grande quantidade em todas as clulas (entre 1.000 e 10.000
molculas de mtDNA por clula), o que lhe confere um menor risco de degradao em
relao ao DNA nuclear. Por estar protegido por uma membrana (da mitocndria), torna-o
extremamente importante no estudo de espcies biolgicas antigas, cujo DNA nuclear, em
geral, se encontra degradado.
DNA mitocondrial
Traduo
Estrutura dos ribossomas, RNAs e protenas ribossomais
Ribossomas: os ribossomas so unidades ultraestruturais onde se sintetizam as protenas,
sendo cada ribossoma constitudo por duas subunidades independentes, designadas por
pequena subunidade ribossmica e grande subunidade ribossmica, que, no seu estado
activo se apresentam unidas e no estado inactivo se encontram livres na matriz
citoplasmtica. Nas clulas eucariticas, a pequena subunidade quimicamente formada por
uma molcula de RNA de 18S e por cerca de 30 protenas diferentes, constituindo, no seu
conjunto, uma estrutura ribonucleoproteica com coeficiente de sedimentao de 40S. A
grande subunidade ribossomal tambm quimicamente uma RNP, com um coeficiente de
sedimentao de 60S. Esta ltima formada por 3 molculas de rRNA: uma de rRNA de
Sebenta de Gentica
FCML
39
28S, outra de rRNA de 5.8S e ainda outra de 5S e por cerca de 50 protenas diferentes. No
seu conjunto molecular cada ribossoma constitui uma RNP de 80S.
No citoplasma das diferentes clulas existem duas populaes de ribossomas. Uma destas
populaes constituda por aglomerados de ribossomas que se encontram aderentes
poro externa da membrana do retculo endoplasmtico rugoso (RER) e interligados por
uma molcula de mRNA. Estas estruturas so responsveis pela sntese e translocao das
protenas para a cisterna do RER. A outra populao constituda por grupos de ribossomas
livres no citoplasma (no associadas a qualquer estrutura membranar). Estes ribossomas
livres, em certas condies fisiolgicas, podem interligar-se por meio de uma molcula de
mRNA, constituindo um polirribossoma (ou polissoma). Os polirribossomas livres sintetizam
protenas livres citoplasmticas, como a hemoglobina, bem como protenas destinadas
importao por parte do ncleo. Estas ltimas atravessam os poros nucleares e vo para o
nucleoplasma, incorporando-se no esqueleto do DNA e dos RNA, constituindo as protenas
nucleares e nucleolares, sendo parte integrante da cromatina, e dos nuclelos. Alm disso,
muitas dessas protenas, como enzimas, interferem nas snteses dos cidos nucleicos.
As duas populaes de ribossomas, livres ou associadas ao RER, so idnticas e
apresentam a mesma morfologia e a mesma composio qumica. Apesar da complexidade
ultraestrutural e molecular do ribossoma, possvel constatar a presena de trs zonas
especficas de ligao para as molculas de RNA, localizadas na pequena unidade
ribossomal, a que foram dados os nome de "centros de ligao" (tambm chamados locais ou
sites). Um centro para o mRNA, que corresponde zona ribossomal em que o mRNA penetra
no ribossoma e por onde este se desloca ao longo do mensageiro. Os outros dois centros
recebem os tRNA e localizam-se em zonas bem distintas na pequena subunidade
ribossomal. Um deles chamado o centro P ou centro peptidil, responsvel pela recepo da
molcula de tRNA que se liga extremidade da cadeia polipeptdica em crescimento. O
outro, chamado centro A ou centro aminoacil, recebe a molcula de tRNA que contm o
aminocido correspondente ao codo do mRNA. Alm dos ribossomas citoplasmticos das
clulas eucariticas, existem ribossomas nos procariontes (mais pequenos). Por outro lado, a
matriz das mitocndrias contm tambm um sistema gentico completo, do qual fazem parte
numerosos ribossomas mitocondriais (tambm chamados mitorribossomas), contendo um
conjunto de rRNA semelhante aos encontrados nos ribossomas dos procariontes.
Tipos de RNA ribossmicos: A sntese do rRNA resulta dum complexo mecanismo em que a
RNA polimerase I se liga sequncia nucleotdica promotora do DNA, situada a montante do
stio de iniciao da sntese e em interaco com factores de iniciao. Na etapa de iniciao,
as duas cadeias do rRNA so separadas por aco da topoisomerase I. O rDNA, nesta zona
activada, perde a sua organizao nucleossmica. O complexo desloca-se ao longo da
Sebenta de Gentica
FCML
40
molcula do rDNA, havendo libertao do factor de iniciao e entrada dos factores de
elongao. A sntese de rRNA ocorre por cpia duma s das duas cadeias do DNAr, no
sentido 5'P - 3' OH. Quando a RNA polimerase I e o factor de elongao atingem a sequncia
nucleotdica correspondente ao stio de terminao, libertam-se da cadeia do rDNA
originando a terminao da sntese. O rRNA transcrito, rRNA de 45S, libertado,
acumulando-se no nuclolo at se iniciar o processo de maturao. O produto primrio da
transcrio uma sequncia nucleotdica de rRNA de 45S (com cerca de 37.700 nucletidos)
que, associada a protenas, forma uma ribonucleoprotena de 80S (RNP 80S). O rRNA 45S
comporta as sequncias do rRNA de 18S, 5.8S e 28S e tambm as sequncias nucleotdicas
intercaladas, que so degradadas no ncleo durante o processo de maturao.
Activao dos aminocidos
During amino acid activation the amino acids (aa) are attached to their corresponding tRNA.
The coupling reactions are catalysed by a group of enzymes called aminoacyl-tRNA
synthetases (named after the reaction product aminoacyl-tRNA or aa-tRNA).
Amino-acil-tRNA sintetases
As molculas de tRNA podem servir de transportadores de aminocidos, estabelecendo uma
ligao covalente entre o grupo hidroxilo da ribose, ligada adenina no terminal 3' e o grupo
carboxilo do aminocido a ser transportado. A reaco catalisada pela enzima amino-acil-
tRNA-sintetase, com dispndio de energia metablica sob a forma de ATP e permite a
esterificao dos grupos OH nas posies 3' ou 2'. A reaco ocorre em duas etapas:
primeiro d-se a formao dum amino-acil-adenilato, com libertao de pirofosfato e depois a
reaco com o tRNA para a formao do amino-acil-tRNA. O aminocido une-se ao
penltimo nucletido no 3end do tRNA atravs de uma ligao ster. Esta ligao conserva
uma considervel poro de energia que vai permitir a formao de uma ligao peptdica
durante a traduo. Um tRNA ligado ao seu aminocido designado charged RNA. A
activao do tRNA depende da presena do aminocido, de ATP e de IF-2 (initiaton factor). A
cada codo do mRNA corresponde um anti-codo dum tRNA, transportador de um
aminocido. Apesar das clulas utilizarem apenas 20 aminocidos, o nmero de sintetases
de aminoacil-tRNA muito superior (61), dependendo do anticodo do tRNA correspondente
ao aminocido, dado que o cdigo degenerado. Na biossntese das protenas, a seleco
do aminocido depende da interaco entre as bases do codo do mRNA e o anticodo do
tRNA, no havendo participao do grupo aminoacil.
Sebenta de Gentica
FCML
41
Iniciao
A iniciao da traduo do mRNA requer a presena de um codo determinado, identificado
como AUG, codo este que codifica para a metionina. Nos eucariontes, a ligao dos
ribossomas ao mRNA realizada inicialmente pela interaco da subunidade 40S com o
mRNA e com a participao de factores proteicos, designados factores de iniciao. Foram
identificados para os eucariontes mais de 10 factores, designados e-IFn. A energia
fornecida pela hidrlise do GTP. Vrias evidncias experimentais sugerem que a seleco do
primeiro codo AUG mediada pela interaco entre a subunidade 40S e os factores IF-4F e
IF-3, associado estrutura cap no terminal 5' do mRNA, de modo que o primeiro AUG
posicionado no centro P, para incio da sntese polipeptdica. Os factores de iniciao, IF-1 e
IF-3 estabilizam a subunidade 40S do ribossoma.O complexo formado pelo mRNA ligado
subunidade 40S e ao primeiro amino-acil-tRNA (contendo metionina, pois o primeiro codo
o AUG) liga-se subunidade 60S, ficando o ribossoma completo com o primeiro aminocido
no stio peptidil (P).
Elongao. Peptidil-transferase e translocao.
A elongao da cadeia polipeptdica, ou seja, a sntese da ligao peptdica entre
aminocidos ordenados segundo a informao transmitida pelo mRNA, ocorre nos
ribossomas em 3 fases de um ciclo que se repete continuamente. Este ciclo, que permite a
ligao de cerca de 40 resduos por segundo, envolve a participao de vrios factores
proteicos, os factores de elongao: EF-1 e EF-2, nos eucariontes. Inicialmente requer a
presena de GTP e de EF-1, que activam o amino-acil-tRNA e induzem a sua ligao ao
codo correspondente no local amino-acil (A) do ribossoma. Aps esta ligao, a enzima
peptidil transferase promove a ligao peptdica entre o novo aminocido e o anterior. Esta
ligao ocorre no local aminoacil (A) do ribossoma. O tRNA do local peptidil fica livre e liberta-
se do ribossoma. Em seguida, na presena de GTP e de EF-2, ocorre a translocao, ou
seja, o movimento do ribossoma, que avana um tripleto no mRNA. Deste modo, o tRNA com
a cadeia peptdica passa para o local peptidil, deixando o amino-acil livre para a entrada de
novo tRNA-amino-acil. Esta translocao ocorre ao longo de todo o mRNA, desde o local de
iniciao AUG at terminao, avanando sempre um tripleto no sentido 5'3'.
Terminao
A terminao, que ocorre quando reconhecido um codo de terminao (UAG, UAA e UGA
- sem tRNA correspondente), requer um s factor de dissociao RF que se liga ao
ribossoma juntamente com GTP. A ligao do factor RF ao codo de terminao induz a
hidrlise da ligao do polipptido ao tRNA que se encontra no local P (atravs da aco da
Sebenta de Gentica
FCML
42
peptidil transferase). Este tRNA dissocia-se do ribossoma com hidrlise de GTP e,
subsequentemente, tambm o mRNA se dissocia do ribossoma, resultando num ribossoma
80S inactivo e pronto a participar novamente na sntese proteica. Vrios ribossomas podem
traduzir uma molcula de mRNA simultaneamente, formando um polirribossoma. Os
ribossomas e polissomas activos podem encontrar-se livres no citoplasma ou associados ao
retculo endoplasmtico. Muitos dos polipptidos neossintetizados no possuem actividade
biolgica e necessitam de sofrer alteraes estruturais para adquirirem a sua funo. As
modificaes ps-traduo de protenas esto dependentes da localizao intracelular final
do polipptido activo. Podemos distinguir vrias modificaes: metilaes, hidroxilaes,
fosforilaes de resduos de aminocidos, adio de resduos glucdicos e remoo de
sequncias peptdicas. O processamento proteoltico ocorre principalmente nas protenas
destinadas a organitos celulares (mitocndrias, lisossomas, retculo endoplasmtico) e em
protenas destinadas a serem secretadas pela clula.
Pptido sinal e topognese
Topognese: The process whereby newly-synthesized proteins are addressed to their correct
locations in the eukaryotic cell has been termed protein topogenesis. Proteins are targeted by
hydrophobic signal sequences to the ER membrane where the Sec61/SecY translocon
mediates the passage of hydrophilic sequences across the membrane and the integration of
apolar transmembrane segments into the lipid bilayer. In this process, the topology of the
polypeptide chain, i.e. the orientation of the signal and subsequent transmembrane segments
in the membrane, is determined.
Pptido sinal: sinal dado por um pptido que permite que as protenas atravessem a
membrana; quando o pptido-sinal no necessrio liberta-se e fica na membrana. Ocorre
em protenas de membrana ou protenas destinadas secreo.
Regies no traduzidas 5UTR e 3UTR
O RNA no comea a ser traduzido logo desde a extremidade 5, pois essa extremidade
apresente o 5UTR (zona no traduzida) que permite a ligao aos ribossomas, logo, estas
zonas no conseguem ser transcritas zona de reconhecimento dos ribossomas. Na zona 3
tambm existe uma regio no traduzida, 3UTR (zona de regulao), sendo que o codo
STOP est antes desta extremidade.
A funo destas regies no traduzidas a de auxiliar a estabilizar o mRNA nos ribossomas.
Sebenta de Gentica
FCML
43
4. Regulao da expresso gentica____________________________
Nveis de regulao da expresso gentica
Fig. 18: Nveis de regulao da expresso gnica
R
R
R
e
e
e
g
g
g
u
u
u
l
l
l
a
a
a
o
o
o
T
T
T
r
r
r
a
a
a
n
n
n
s
s
s
c
c
c
r
r
r
i
i
i
p
p
p
c
c
c
i
i
i
o
o
o
n
n
n
a
a
a
l
l
l
Remodelao da cromatina
(Acetilao e Metilao de CG) e consequente alterao da estrutura da cromatina- so
modificaes epigenticas j que no se baseiam na alterao da sequncia do DNA.
ACETILAO: promove transcrio.
O relaxamento da cromatina provocado por acetilao das histonas: adio de
grupos acetil.
A acetilao das histonas pela histona-acetiltransferase (HATs) recrura outros
factores de remodelao como SWI/SNF, o que facilita a formao do complexo de
iniciao com ligao de factores de transcrio (TF) e da RNApil.
A remodelao da cromatina pode ser afectada pela fase do ciclo celular.
Os requisitos de remodelao podem ser especficos de gene.
Sebenta de Gentica
FCML
44
METILAO: actua de modo a silenciar os genes, isto , grandes nveis de metilao esto
associados a genes que no so activamente transcritos (determinados genes controlam a
metilao do DNA, por meio de metilases, como por exemplo, o gene xist). Os locais CpG
so os mais metilados. O DNA de cada indivduo possui um determinado nmero de bases
metiladas, que permite o reconhecimento do padro de metilao como prprio da clula e a
sua auto-regulao mecanismo de autorreconhecimento e autorregulao. Nos mamferos,
2 a 7% das bases C so modificadas pela aco de uma enzima, que cria 5'-metil-citosina
pela adio de grupos metilo base. Quando um dinucletido CpG, que se encontra metilado
em ambas cadeias, sofre replicao, o resultado so duas molculas filhas, em que cada
uma contm uma cadeia metilada e outra normal. A DNA metilase reconhece a semi-
metilao destas molculas e metila as citosinas das cadeias filhas. Assim, a maior parte das
5'-metil-citosinas encontram-se nas sequncias 5'-CpG-3', embora nveis mais reduzidos de
metilao tambm possam ser encontradas noutras sequncias.
NOTA: Quanto alterao da estrutura da cromatina, deve-se ter presente o efeito da
poli-ADP ribosilao das histonas. As protenas histnicas H1, H2B e lmina B, uma vez
ribosiladas, perdem a afinidade para o DNA, devido carga altamente negativa dos
polmeros de ADP-ribose. Assim, este processo pode constituir uma forma relativamente
simples de desfazer a ordem mais elevada da organizao da cromatina, de modo a facilitar
o acesso dos factores de transcrio s sequncias reguladoras da expresso gnica.
F Fa ac ct to or re es s d de e t tr ra an ns sc cr ri i o o, , p pr ro om mo ot to or re es s, , e en nh ha an nc ce er rs s e e s si il le en nc ce er rs s
Os promotores, enhancers e silenciadore so elementos regulatrios em cis: a protena vem
da mesma sequncia do gene.
Os promotores so combinaes de curtos elementos de sequncia, normalmente
localizados na regio imediatamente a montante do gene e servem para iniciar a transcrio.
Podem ser subdivididos em diferentes componentes:
- Regio promotora central: contm componentes que direccionam o complexo de transcrio
basal para iniciar a transcrio de genes e que, na ausncia de elementos reguladores
adicionais, permite a expresso constitutiva do gene mas em nveis muito baixos (basais).
Elementos promotores centrais esto tipicamente localizados muito prximos do stio de
iniciao da transcrio e incluem: TATA box (circundado por sequncias ricas em GC) e
reconhecido pela protena de ligao ao TATA box, subunidade de TFIID); a sequncia BRE
(TFIIB Recognition Element) est localizada a montante do elemento TATA e reconhecida
pelo componente TFIIB; a sequncia INR (iniciadora) localizada no stio de iniciao da
transcrio e o DPE (Downstream Promoter Element) elemento promotor a jusante.
Sebenta de Gentica
FCML
45
- Elementos promotores no centrais: podem ser encontrados na regio promotora central
mas esto tipicamente localizados na sequncia a montante dessa. Esta regio algumas
vezes referida como regio promotora proximal (em oposio regio promotora distal,
usada para descrever regies ainda mais distantes, a montante). Existem normalmente
mltiplos stios de reconhecimento para alguns factores de transcrio ubquos sequncia-
especficos tal como GC-box e CCAAT-box (ambas so sequncias reforadores- enhancers)
Reforadores (enhancers): constituem um grupo de elementos cis-atenuantes, de
sequncias curtas que intensificam a actividade de transcrio basal de genes especficos
dos eucariotas por meio dos promotores centrais. Ao contrrio dos promotores, cujas
posies relativas ao local de incio da transcrio so relativamente constantes, os
reforadores localizam-se a distncias variveis e frequentemente grandes desse stio e a
sua funo concreta depende da sua orientao. Pode localizar-se antes da sequncia
promotora, a seguir ao gene (no sentido em que se processa a transcrio), interposta no
prprio gene ou ainda na cadeia complementar de DNA do gene em causa.
Silenciadores: servem para reduzir os nveis de transcrio. Temos silenciadores
clssicos- elementos independentes da posio que orientam um mecanismo activo de
represso da transcrio- e elementos reguladores negativos- dependentes da posio
resultando num mecanismo de represso passivo.
Insuladores (elementos de fronteira): regies de DNA funcionam como bloqueadores da
propagao da influncia de agentes que possuem um efeito positivo sobre a transcrio
(reforadores) ou negativo (sileciadores).
Elementos de Resposta: funcionam para modular a transcrio em resposta a um
estmulo externo especfico (hormonas normalmente)
Promotores
centrais
Elementos
resposta
Silenciador
Reforadores
Sebenta de Gentica
FCML
46
Factores de transcrio
So elementos regulatrios em trans: so motivos de protenas de ligao ao DNA
Nos eucariotas, a classe de factores de transcrio mais importante a general transcription
factors (GTFs). A maioria no se liga directamente ao DNA mas une-se RNA polimerase
formando um complexo pre-iniciao que se liga ao promotor. Os GTFs mais comuns so:
TFIIA, TFIIB, TFIID, TFIIE, TFIIF e TFIIH.
Os factores de transcrio contm os seguintes domnios:
1) DNA binding domain (DBD): liga-se a sequncias de DNA especficas
Sequncias em hlice (HTH helix-turn-helix) 3 ou 4 hlices, em que uma de
reconhecimento de DNA. Actuam em dmeros que formam um ngulo (turn).
Zn finger forma quelatos que apresentam dedos de ligao de Zn (a Cys e a His
contribuem para a formao do quelato). Possuem 2 -hlices unidas por uma estrutura em
folha pregueada que coordenam ies de Zn (atravs de cistenas e histidinas).
-hlice loop (HLH) pedao solto de protena que constitui uma ansa descada
Leucina zipper -helix sequencias de leucinas (5 leucinas espaadas por 7 aminocidos)
em monmeros que estabelecem interaces hidrofbicas para formar dmeros proteicos de
reconhecimento de DNA. Heterodmero que se liga ao DNA. Encaixa como um fecho clair
com outra protena que tambm possua leucina, formando homodmeros (protenas iguais)
ou heterodmeros (protenas diferentes), determinando o silenciamento ou traduo de um
gene.
Fig. 19: HTH, HLH, Zn finger, Leucine zipper -helix
determinam a capacidade de uma protena se ligar ao DNA
Sebenta de Gentica
FCML
47
2) Trans-activating domain (TAD): contm binding sites (activation functions-AFs) para outras
protenas, como co-reguladores de transcrio.
3) Single sensing domain (SSD): opcional. Ex: ligand binding domain.
R Re eg gu ul la a o o n no o p pr ro oc ce es ss sa am me en nt to o d de e m mR RN NA A e e m mo od di if fi ic ca a e es s p p s s- -t tr ra an ns sc cr ri ip pc ci io on na ai is s
A forma como se opera o processamento do RNAhn (RNA heterogneo ou RNA pr-
mensageiro) tambm se reflecte na regulao da expresso gnica, em relao com a
degradao, a poliadenilao, o splicing e o editing de RNA (ver pgina 25). Nas clulas dos
mamferos, os candidatos a reguladores do splicing so a famlia SR de protenas de ligao
a RNA (que possuem um domnio C-terminal distinto e rico em dipeptdeos serina-arginina) e
algumas protenas HnRNP (heterogeneous nuclear ribonucleoprotein particle). Essas
protenas so conhecidas por promoverem vrias etapas da montagem do spliceossomo e
por se ligarem a sequncias reforadoras de splicing, sequncias reguladoras que podem
aumentar o reconhecimento do stio de splicing.
NOTA: Protena Slo protena da cclea que forma protenas diferentes que contribuem para
a selectividade da frequncia coclear: canais de K
+
activados por Ca
2+
e por despolarizao
da membrana. O splicing alternativo de slo gera canais de K
+
activados por Ca
2+
com
cinticas distintas.
R Re eg gu ul la a o o p p s s- -t tr ra an ns sc cr ri i o o
Para alm da regulao no processamento de mRNA, a regulao ps-transcripcional inclui:
a) regulao dos ncRNAs (ver pgina 34)
b) non-sense mediated RNA decay
- devido a uma mutao pode ocorrer que o codo STOP aparea um bocadinho mais cedo
no afecta grandemento o organismo.
- Se o codo estiver a meio caminho o pptido resultante pode provocar patologias.
non-sense mediated RNA decay reconhece o STOP mal localizado, onde se agrega uma
srie de protenas e levam a um decay e a um recycling das protenas. O reconhecimento por
estas protenas pode ocorrer no ncleo.
Mutaes na Distrofina relacionada com o sistema mRNA decay; provoca distrofias
musculares por falta de distrofina. A distrofia pode surgir pela doena de Duchene ou pela
Doena de Becker (mais leve).
Sebenta de Gentica
FCML
48
R Re eg gu ul la a o o e e o or rg ga an ni iz za a o o d da a c cr ro om ma at ti in na a
Ver NOTA da regulao transcripcional a)
R Re eg gu ul la a o o a a n n v ve el l d da a t tr ra ad du u o o
A eficcia da traduo do mRNA influenciada pela acessibilidade do codo AUG ao
ribossoma, a disponibilidade dos ribossomas e do tRNA, a estabilidade do mRNA e a
velocidade da sntese de cada molcula polipeptdica. O controlo da expresso gnica pela
estabilidade do mRNA foi verificado no processo de sntese da tubulina, em que se observou
uma diminuio do nmero de molculas de mRNA em correlao com um aumento do
nmero de dmeros de tubulina. A fosforilao da protena EIF-2, ao traduzir-se na represso
do incio da traduo contitui uma das formas de regulao a este nvel. Na verdade, na
condio fosforilada, no ocorre a ligao de uma molcula de GTP molcula EIF-2, uma
condio necessria para que esta molcula providencie o transporte do tRNA iniciador para
a unidade menor do ribossoma.
Regulao de traduo de mRNA com cap: IRES ajuda a decidir se aceita protenas
com cap ou sem cap.
Fig.22: Regulao de traduo de mRNA com cap
R Re eg gu ul la a o o p p s s- -t tr ra ad du uc ci io on na al l
a) Clivagem por proteases
Um polipeptdeo pode no ser funcional na forma em que produzido a nivel ribossomico
como ocorre com o pepsinogenio ou a pr-insulina sendo necessria a sua clivagem
Sebenta de Gentica
FCML
49
prvia para que a molcula possa actuar. Por outro lado a clivagem depende de enzimas
presentes no ambiente em que se encontra o produto. A modificao da estrutura de um
polipeptideo pode ainda ocorrer por aco de proteases que levem sua ciso em
diferentes locais, de modo a originar molculas com actividade diferente em funo do
local em que se opera a ciso.
b) Disponibilidade de ies
Caso da hemoglobina em que pode haver uma deficincia devida a falta de ferro, para
uma capacidade normal de sntese proteca.
c) Organizao em oligmeros
O produto codificado por um gene pode actuar de uma forma monomrica ou associar-se
a outras moleculas polipeptidicas iguais ou diferentes. Nos casos em que se formam
oligmeros, embora haja a produo inicial de um nmero equivalente de monmeros
normais e de monmeros com mutao ainda que no dominante, a capacidade funcional
dos oligmeros que incorporam monmeros mutados afectada, havendo uma perda de
funo bastante mais acentuada do que se observa para idnticas condies de
heterozigotia, quando a molcula funcional um monmero
d) Fosforilao (regulao de actividade enzimtica; cascatas de transduo de sinal)
O grau de fosforilao das protenas, a nvel da serina, treonina e tirosina, pode
influenciar a sua actividade, como se verifica com a protena p105RB codificada pelo
gene RB. Trata-se de uma forma de regulao fina, mediada por um balano entre a
actividade das proteinacinases (envolvidas na fosforilao) e a actividade das fosfatases
(com capacidade para removerem os fosfatos). A fosforilao da protena inibe a
formao de complexos com protenas especficas, deixando-as livres. Tratando-se da
complexao de factores de transcrio como a protena E2F, a manuteno na forma
livre vai possibilitar a sua ligao regio reguladora dos genes alvo e a promoo da
sua expresso. Pelo contrrio, no estado no fosforilado, observa-se a formao de
complexos com a protena alvo que inibe a sua aco.
e) Glicosilao (no aparelho de Golgi; fulcral para as protenas de exportao)
f) Acetilao; Metilao; Hidroxilao; Ribosilao; Pontes dissulfeto
Sebenta de Gentica
FCML
50
g) Ubiquitinao e degradao
Ubiquitina:
- protena presente em todas as clulas eucariotas.
- Contm uma sequncia altamente conservada, idntica no Homem, peixes e insectos
(entre levedura e Homem s diferem 3 dos 76 aminocidos).
- Participa em diversas funes celulares, como degradao proteica, estrutura da
cromatina e shock trmico.
- Conjuga-se com protenas pela extremidade COOH.
Fig. 23: Ubiquitinao
R Re eg gu ul la a o o e ep pi ig ge en n t ti ic ca a: : modificaes hereditrias que no dependem de mudanas na
sequncia de DNA so denominadas epigenticas.
Sebenta de Gentica
FCML
51
5. Gentica de Populaes_____________________________________
Population genetics is a useful tool for studying evolution and quantifying how evolution
operates. It emphasizes counting allele and genotype frequencies to understand how
phenotype frequencies in a population change over time. Models can be built that incorporate
the varied effects of selection, genetic drift, migration, etc. For example, with population
genetics, you can ask: How long would it take for a given allele to be fixed given a certain
selective force for it?
A uniformidade e a disjuno mendeliana dos caracteres.
Dominncia e recessividade.
Em gentica, um trao dominante aquele que expresso fenotipicamente em heterozigotos
enquanto que um recessivo s expresso em homizogotos.
Conceito de locus e de alelo.
Locus: localizao cromossmica nica que define a posio de um gene individual ou de
uma sequncia de DNA
Alelo: uma das vrias formas alternativas de um gene ou sequncia de DNA numa posio
especfica do cromossoma (lcus). Em cada lcus autossmico cada indivduo possu dois
alelos, um materno e outro paterno
Heterogeneidade gentica e allica.
Heterogeneidade gentica: Quando um distrbio gentico, que parece ser uma entidade
nica analisado descobre-se com frequncia que geneticamente heterogneo, ou seja,
que inclui diversos fentipos que so semelhantes mas determinados por gentipos
diferentes. A heterogeneidade pode resultar de mutaes distintas no mesmo locus
(heterogeneidade allica), mutaes em loci diferentes (heterogeneidade no allica), ou
ambas. O reconhecimento da heterogeneidade gentica um aspecto importante do
diagnstico clnico. Um carcter dito geneticamente heterogneo quando mutaes em
diferentes loci ou diferentes mutaes no mesmo loci podem conduzir, independentemente,
ao mesmo carcter ou caracteres, que so clinicamente difceis de distinguir. Por outras
palavras, fala-se em heterogeneidade gentica quando, para um mesmo fentipo, podem
existir vrios gentipos. Exemplos: Tay-Sachs (mesmo locus - allica); Charcot-Mane-Tooth
(diferentes loc i- locus).
Heterogeneidade allica: Condio em que um fentipo ou fentipos muito semelhantes so
determinados, de um modo independente, por alelos diferentes localizados no mesmo locus.
Sebenta de Gentica
FCML
52
Fig. 24: Legenda:
(A) Autossmica dominante;
(B) Autossmica recessiva;
(C) Ligada ao X recessiva;
(D) Ligada ao X dominante;
(E) Ligada Y
O risco para os indivduos marcados : (A) 1 em 2 (B) 1 em 4 (C) 1 em 2 homens ou 1 em 4 do total de
descendentes (D) 100% para mulheres e muito baixo para os homens (praticamente negligencivel)
Frequentemente, uma patologia pode ser provocada por diferentes tipos de mutaes num
gene heterogeneidade allica
A segregao mendeliana dos caracteres
Sebenta de Gentica
FCML
53
Fig. 25: Complicaes dos padres mendelianos bsicos:
(A) Recessivo comum tal como grupo sanguneo O, pode aparentar um padro dominante (B) Autossmica
dominante com no penetrncia em II2. (C) Autossmica dominante com expresso varivel: nesta famlia com o
sndrome de Waardenburg, 1 quadrante preenchido = perda de audio; 2 quadrante = colorao de olhos
diferente; 3 quadrante = madeixa branca; 4 quadrante = cabelos brancos prematuros. (D) Imprinting gentico:
nesta famlia o paraganglinoma autossmico dominante s se manifesta quando o gene herdado do pai. (E)
Imprinting gentico: o syndrome de Beckwith-Wiedemann, autossmico dominante nesta famlia s se manifesta
quando o gene herdado da me. (F) Incontinentia pigmenti ligada ao X dominante. Homens afectados
resultaram em abortos espontneos (quadrado pequeno). (G) Ligada ao X recessive em que o inbreeding origina
uma mulher afectada e aparenta transmisso de homem para homem. (H) Uma nova mutao autossmica
dominante aparentando um padro autossmico ou ligado ao X recessivo.
Recombinao por crossing-over e o fenmeno de linkage
Recombinao por crossing-over: Designa-se por recombinao a formao de novas
combinaes entre os genes. uma troca de patrimnio gentico entre loci de cromossomas.
O crossing-over um fenmeno que ocorre durante a meiose (profase I), havendo troca de
segmentos de cromatdios entre cromossomas homlogos, promovendo assim uma maior
diversidade na gerao seguinte. Quanto mais afastadas estiverem duas pores de DNA,
Sebenta de Gentica
FCML
54
maior ser a probabilidade de ocorrer recombinao por crossing-over durante a meiose. O
centiMorgan (cM), unidade de comprimento do DNA , sobretudo, uma unidade de
recombinao, pois corresponde probabilidade de dois genes se separarem durante a
meiose (cerca de 1% para um afastamento de 10
6
pares de bases). Uma causa frequente de
mutao envolve a deleco ou duplicao mediada por recombinao entre sequncias de
DNA altamente semelhantes ou idnticas. A recombinao que ocorre entre cromossomas ou
cromtides irms mal emparelhadas pode levar deleco ou duplicao de genes. Acredita-
se, portanto, que o mecanismo de crossing-over desigual, ou seja, entre sequncias de DNA
semelhantes que esto desalinhadas seja responsvel por diversas variantes genticas (por
exemplo, a Hemoglobina de Lepore).
Fig. 26: Recombinao por crossing-over
Fenmeno de linkage:
O fenmeno de linkage ou ligao gnica a tendncia de genes ou outras sequncias de
DNA em lcus especficos serem herdados juntos como consequncia da sua proximidade
fsica num nico cromossoma. A probabilidade de transmisso de dois genes juntos depende
da frequncia com que so separados por crossing-over, estando esta relacionada com a
distncia que os separa. Assim, quanto mais prximos esto os genes num cromossoma
menor ser a probabilidade de crossing-over durante a meiose (95% de probabilidade de no
ocorrer crossing-over quando os genes esto separados por cerca de 5 milhes de bases).
Se os genes esto separados por 5cM, a probabilidade de serem herdados juntos de 95%.
Quando dois pares de genes alelos esto situados (num par de homlogos) de tal forma que
no ocorre troca/recombinao entre eles, diz-se que h linkage total. Caso haja
troca/recombinao, o linkage parcial. Para estudos de ligao gnica (linkage) podem ser
usados os RFLPs, os VNTRs e os SNPs.
O LOD score (logarithm of the odd) usado na avaliao de genealogias quanto ligao.
um mtodo estatstico para verificar se dois loci esto em ligao (linkage). uma funo da
Sebenta de Gentica
FCML
55
fraco de recombinao (u) caracterizada pelo logaritmo do quociente da probabilidade de
linkage sobre a de no linkage. Num conjunto de famlias, a probabilidade total de linkage o
produto das probabilidades de cada famlia individualmente. Sendo logaritmos, os LOD
scores das vrias famlias podem ser somados
Transmisso matrilinear do DNA mitocondrial.
O DNA mitocondrial (mtDNA) apresenta-se sob a forma de um cromossoma circular. H
vrias cpias deste cromossoma circular por mitocndria e milhares por clula. Determinou-
se a sequncia completa do mtDNA e sabe-se que codifica 2 tipos de RNA ribossmico, 22
RNA de transferncia e 13 polipptidos que so subunidades de enzimas da fosforilao
oxidativa. O mtDNA replica-se dentro da mitocndria e a mitocndria divide-se por fisso
simples. Durante a citocinese, as mitocndrias distribuem-se aleatoriamente pelas clulas
filhas. Quando uma clula, contendo mtDNA normal e mutante, se divide, as suas clulas
filhas podem conter apenas mtDNA normal, apenas mtDNA mutante ou ambos. Como a
expresso fenotpica de uma mutao no DNA mitocondrial depende das propores
relativas de mtDNA normal e mutante, a variabilidade de expresso uma caracterstica dos
heredogramas dos distrbios mitocondriais. Actualmente, conhecem-se vrias doenas
neuromusculares associadas a mutaes do mtDNA, nomeadamente a doena de Leber
(neuropatia ptica hereditria de Leber - LOHN), que se expressa fenotipicamente, com a
perda bilateral rpida da viso central, em virtude da morte do nervo ptico em adultos
jovens. Uma caracterstica particular do mtDNA a sua herana materna ou matrilinear. Ao
contrrio do espermatozide, o ovcito rico em mitocndrias sendo, portanto, a me que
transmite o seu mtDNA a toda a descendncia. Outro factor importante a heteroplasmia
caracterstica das doenas associadas s mitocndrias. Cada clula humana contm vrios
genomas mitocondriais. Em alguns pacientes com doenas mitocondriais, cada genoma
mitocondrial apresenta a mutao causadora da doena (homoplasmia) mas noutros casos,
em cada clula existe uma populao mista de genomas normais e de genomas mutantes
(heteroplasmia). Diversa do mosaicismo gentico nuclear, que tem origem ps-zigtica, a
heteroplasmia pode ser transmitida de uma me heteroplsmica para uma crianas
heteroplsmica.
Gentica formal.
F Fr re eq qu u n nc ci ia a g g n ni ic ca a, , f fr re eq qu u n nc ci ia a g ge en no ot t p pi ic ca a
Frequncia genotpica: frequncia de determinado gentipo na populao
Sebenta de Gentica
FCML
56
Frequncia gnica: numa populao podem existir diferentes alelos de um determinado lcus
embora cada indivduo tenha apenas dois desses alelos os quais podem ser identicos ou
diferentes. Podemos imaginar um conjunto gnico constitudo por todos os alelos do lcus A
de uma populao. A frequncia gnica do alelo A
1
a proporo de todos os alelos A do
conjunto gnico que so A
1
. Se retirarmos um alelo ao calhas do gene pool, a probabilidade
de ser A
1
p e a de ser A
2
q sendo que p e q variam entre 0 e 1 e p+q=1.
Frequncias genotpicas em equilibrio Hardy- Weinberg para as frequncias allicas p (A) e q
(a):
Nota: estas frequncias sero observadas mesmo que A e a no sejam os nicos alelos do
lucs (heterogeneidade allica)
Exemplo: Fibrose qustica (a mais frequente situao recessiva)- incidncia: cerca de 1/1600
nascimentos
Qual a frequncia de portadores saudveis ?
q2 = 1/1600
q = \1/1600 = 1/40
2pq = 2(39/40 x 1/40)= 1/20
Fig. 27: HardyWeinberg principle for two alleles: the horizontal axis shows the two allele frequencies p and q and
the vertical axis shows the genotype frequencies. Each graph shows one of the three possible genotypes.
Sebenta de Gentica
FCML
57
E Eq qu ui il l b br ri io o d de e H Ha ar rd dy y- -W We ei in nb be er rg g
A lei de Hardy-Weinberg enuncia que existe um grupo de genes que, numa determinada
populao, esto em equilbrio - "Em populaes muito grandes, em que os cruzamentos
ocorrem ao acaso e no sujeitos aco de factores de evoluo, a frequncia de cada alelo
tende a manter-se constante, de gerao em gerao." Os gentipos distribuem-se
proporcionalmente s frequncias dos alelos numa populao, permanecendo constantes de
gerao em gerao.
Uma populao que demonstra as caractersticas bsicas da lei de Hardy-Weinberg diz-se
que est em equilbrio de Hardy-Weinberg. No entanto, a lei de Hardy-Weinberg baseia-se
em vrias permissas fundamentais, que nem sempre so verdadeiras para as populaes
humanas reais:
1. Ausncia de mutaes
2. O locus em causa exibe uma taxa de mutao constante e os alelos mutantes
perdidos por morte so substitudos por neomutaes.
Equilbrio seleco-mutao:
Condio autossmica recessiva: = sq
2
ou = F(1-f)
Condio autossmica dominante: = sp ou = F(1-f)
Condio recessiva ligada ao X: = sq/3 ou = 1/3 F(1-f)
Em que:
= taxa de mutao de gene por gerao
p,q= frequncias gnicas
s= coeficiente de seleco
f= adaptao biolgica= 1-s
F= frequncia da condio na populao
3. No existem vantagens selectivas de qualquer fentipo, todos os gentipos num locus
so igualmente viveis.
4. O tamanho da populao nunca inferior a 10 000 indivduos.
5. Ausncia de qualquer alterao da estrutura populacional por migrao, garantindo o
isolamento gentico e a no modificao da frequncia gentica.
6. A populao caracteriza-se pela existncia de panmixia de modo que a probabilidade
de cruzamento seja apenas um reflexo da frequncia gentica da gerao progenitora
paralelismo com o equilbrio de linkage (a probabilidade de 2 genes se associarem em
linkage depende da sua frequncia na populao).
Sebenta de Gentica
FCML
58
Filhos de casais em endogamia:
Frequncia de recessivos (aa): q
2
+ Fq
Frequncia de heterozigotos (Aa): 2pq 2Fpq
Frequncia de homozigotos dominantes (AA): p
2
+ Fpq
(sendo F o quociente de consaguinidade)
Se numa dada populao, uma ou mais destas permissas no for vlida, os gentipos podem
no estar em equlbrio de Hardy-Weinberg.
A principal aplicao da lei no clculo das frequncias gnicas. Assim, podemos determinar
se a transmisso de uma doena dominante ou recessiva comparando a frequncia terica
com a frequncia fenotpica observada.
B Bi ia al le el li ia a e e p po ol li i- -a al le el li ia a
Bialelia: existncia de apenas dois alelos numa populao para um nico locus.
Poli-alelia: existncia de vrios alelos numa populao, que concorrem para um nico locus.
O gentipo de cada indivduo consiste num dos pares de alelos resultantes das combinaes
possveis entre o total de alelos.
P Po ol li im mo or rf fi is sm mo os s
Rigorosamente a existncia de duas ou mais variantes (alelos, fentipos, variantes de
sequncias, variantes de estrutura cromossmica) em frequncias significativas na
populao. A utilizao de uma forma menos rigorosa do termo entre os geneticistas
moleculares inclui: 1) qualquer variante de sequncia presente numa frequncia >1%
2)qualquer variante da sequncia no-patognica independentemente da frequncia.
I Is so ol la ad do os s g ge en n t ti ic co os s
Certas populaes revelam tendncia para a criao de isolados genticos assentes em
bases geogrficas, culturais e religiosas (judeus, ciganos, etc). Estes isolados favorecem a
endogamia, casamentos consanguneos com tendncia para uma igualdade de genes, o que
favorece o aparecimento de gentipos recessivos com alta frequncia. No h vantagem
selectiva para determinadas associaes de genes e as populaes encontram-se em
desequlibrio gentico. Podemos referir a doena de Tay-Sachs, frequente entre os judeus
Ashakenazi da Amrica do Norte. Esta doena corresponde a uma deficincia da enzima
hexosaminidase A que cliva o radical terminal do ganglisido, levando a uma acumulao de
GM2 ao nvel do sistema nervoso central. Estas crianas ficam cegas, apresentam uma
Sebenta de Gentica
FCML
59
regresso mental e fsica, acabando por falecer durante a infncia. A tiroxmia, doena
causada pela incapacidade de degradar a tiroxina, outra situao com frequncia
aumentada em determinados isolados genticos.
P Pa an nm mi ix xi ia a e e e en nd do og ga am mi ia a
Panmixia: acasalamento livre e fortuito, na ausncia de qualquer seleco, partindo da
hiptese de que os cnjuges se escolhem de forma aleatria, que a probabilidade de
encontrar um parceiro igual para todos os indivduos e que a fecundidade igual para todos
os casais e a viabilidade dos descendentes tambm igual. Estas condies so
preenchidas nos grandes grupos da populao humana.
Endogamia: um sistema em que os acasalamentos se do entre indivduos aparentados,
relacionados pela ascendncia, ou seja, a unio de indivduos mais aparentados do que a
mdia da populao. Tem como efeito gentico a diminuio da heterozigotia e o aumento da
homozigotia, e, como efeito fenotpico, uma grande manifestao de genes recessivos, que
acabam por resultar na perda de vigor, assim como a perda da varincia, medida que
aumenta o parentesco.
E Ef fe ei it to o f fu un nd da ad do or r
A frequncia gentica varia de um modo desordenado e imprevisvel - um gene pode atingir
uma frequncia de 0 (o gene foi eliminado da populao) ou de 1 (o gene est presente em
toda a populao). Neste ltimo caso ter ocorrido a fixao que no pode ser revertida. Este
fenmeno aplica-se ao estabelecimento de alelos mutantes novos na populao e formao
de subpopulaes pequenas, por isolamento de uma populao inicial. Na nova populao,
as frequncias genticas podem ser muito diferentes das da populao original porque o
novo grupo representa apenas uma pequena amostra do grupo parental. Se um dos
fundadores do grupo possuir, por acaso, um alelo relativamente raro, esse alelo pode tornar-
se fixo no novo grupo e com uma frequncia relativamente alta. Esta a base do efeito
fundador (que uma causa da deriva gentica). Dois exemplos so: Doena de Machado-
Joseph, na Ilha das Flores e a Polineuropatia amiloidtica familiar, na Pvoa do Varzim (ver
pg. 100).
M Mi ig gr ra a e es s p po op pu ul la ac ci io on na ai is s d de e a al le el lo os s
Os indivduos que se introduzem numa populao relativamente isolada e de nmero limitado
introduzem nela genes que previamente no existiam. De facto a migrao implica o fluxo
gentico que definido como a lenta difuso de genes atravs de uma barreira populacional
correspondendo a um processo que envolve, de um modo geral, o movimento de indivduos
Sebenta de Gentica
FCML
60
de uma populao para outra. Essa migrao provoca uma alterao gradual das frequncias
dos genes, na medida em que uma populao pode receber genes quando h entrada de
indivduos ou perder genes quando h sada (fluxo positivo e fluxo negativo,
respectivamente). Os genes das populaes migrantes incorporam-se gradualmente no pool
gnico da populao para a qual migraram. Com efeito, ocorre miscigenao da populao
migrante com a populao anfitri e, logo, a frequncia dos alelos modifica-se e novas
combinaes genticas so criadas.
Conceito Out of Africa: Researchers compared 650,000 genetic markers in nearly a thousand
individuals from 51 populations around the globean unprecedented level of detail for a
human genetic study and found out that you get less and less variation the further you go from
Africa. Such a pattern fits the theory that the first modern humans settled the world in
stepping-stone fashion after leaving Africa less than 100,000 years ago. As each small group
of people broke away to found a new region, it took only a sample of the parent population's
genetic diversity. If you keep sampling like that, then mathematically you must lose variation.
In particular, the pattern of variation shows that the route of migration out of Africa was into
the Middle East linkageand then to the rest of Eurasia, the Americas, and Oceania. That tends
to agree with what we're seeing on the Y-chromosome side.
C Co on nc ce ei it to o d de e b bo ot tt tl le en ne ec ck k. . E Ex xp pa an ns s o o e e m mi ig gr ra a o o
uma tendncia evolutiva, causa da deriva gentica, onde h uma reduo drstica do
tamanho da populao (normalmente devido a desastres naturais que causam muitas
mortes). totalmente arbitrrio o fentipo que continua a viver (e contribui para futuras
geraes) e o que eliminado.
Fig.28: Efeito Bottleneck
Sebenta de Gentica
FCML
61
D De es se eq qu ui il l b br ri io o d de e l li in nk ka ag ge e e e s su ua a v va ar ri ia a o o a a n n v ve el l p po op pu ul la ac ci io on na al l
Normalmente, os genes ligados (em linkage) deveriam estar em equilbrio, isto , as
propores relativas das possveis combinaes s deveriam ser determinadas pela
frequncia dos alelos dos genes ligados de uma populao - as frequncias gnicas
deveriam corresponder s frequncias fenotpicas: equilbrio de ligao (linkage). Ou seja,
para haver equilbrio no podem ocorrer ligaes preferenciais entre dois alelos sendo estas
dependentes apenas do acaso. No entanto, por vezes ocorrem desequilbrios porque as
ligaes no so feitas ao acaso. Este desequilbrio entre loci diminui ao longo do tempo
como resultado da recombinao. Explicaes possveis, para o desequilbrio de linkage so:
- mutao na populao, que pode ter sido demasiado recente para permitir reestabelecer o
equilbrio;
- inverso cromossmica envolvendo os genes ligados (a recombinao suprimida dentro
da inverso; cromtides com deficincia de duplicao);
- a seleco natural favore combinaes particulares de alelos nos loci ligados (loci HLA).
Sejam p1 e p2 frequncias de 2 alelos em 2 loci em ligao:
a frequncia do haplotipo ser: h = p1 . p2
- Se h p1 . p2 , teremos: = h - p1 . p2 , isto , desequilbrio linkage (D)(D)
0 taxa de 1 u por gerao, de onde : D
g
= (1 u) D
g-1
Graf. 1: Desequilbrio de linkage: em funo do nmero de geraes, influncia de u
H Ha ap pM Ma ap p e e b bl lo oc co os s h ha ap pl lo ot t p pi ic co os s
Blocos haplotpicos: Raramente a recombinao ir separar dois lcus que se situam muito
prximos entre si no mesmo cromossoma porque s um crossing-over situado exactamente
Sebenta de Gentica
FCML
62
no pequeno espao entre eles resultar numa recombinao. Por isso, conjuntos de alelos
situados no mesmo pequeno segmento cromossmico tendem a ser transmitidos em bloco
na genealogia. Um bloco de alelos assim reconhecido como hapltipo. Os hapltipos
marcam segmentos cromossmicos reconhecveis, que podem ser seguidos atravs de
genealogias e de populaes. O bloco haplotpico corresponde a um haplotipo que tende a
repetir-se, ou seja, dois ou mais genes em loci em linkage que tendem a ser segregados em
conjunto. O desequilbrio de linkage explica a formao de blocos haplotpicos.
HapMap:
The HapMap is a catalog of common genetic variants that occur in human beings. It describes
what these variants are, where they occur in our DNA, and how they are distributed among
people within populations and among populations in different parts of the world. The
International HapMap Project is not using the information in the HapMap to establish
connections between particular genetic variants and diseases. Rather, the Project is designed
to provide information that other researchers can use to link genetic variants to the risk for
specific illnesses, which will lead to new methods of preventing, diagnosing, and treating
disease.
A construo de um HapMap ocorre em trs passos (ver figura 29):
(a) SNPs are identified in DNA samples from multiple indivduals
(b) Adjacent SNPs that are inherited together are compiled into "haplotypes."
(c) "Tag" SNPs within haplotypes are identified that uniquely identify those haplotypes. By
genotyping the three tag SNPs shown in this figure, researchers can identify which of the four
haplotypes shown here are present in each individual.
Fig.29: The construction of the HapMap
Sebenta de Gentica
FCML
63
D De er ri iv va a g ge en n t ti ic ca a
Quando ocorre uma mutao, a sua frequncia representada apenas por uma cpia entre
todas as cpias daquele gene na populao. A probabilidade de que esta nova mutao
sobreviva de uma gerao prxima determinada pelo acaso e por seleco natural.
Dependendo do acaso, a frequncia do alelo flutua de gerao em gerao, sobretudo se a
populao for de tamanho reduzido. Este fenmeno denomina-se deriva gentica. A deriva
gentica , portanto, a flutuao da frequncia relativa dos genes ao acaso em pequenas
populaes. Pode favorecer, o estabelecimento de genes que no so favorveis ou at
neutros, mas no realmente prejudicias.
F Fi it tn ne es ss s e e d di is st tr ri ib bu ui i o o g ge eo og gr r f fi ic ca a
Numa populao surgem alelos novos por neomutao e so mantidos ou removidos por
seleco (natural ou artificial). Aps o aparecimento de uma nova mutao, a sua
manuteno na populao depende do fitness (adaptabilidade) dos indivduos que a tm, em
comparao com os indivduos que possuem outros alelos no locus em causa. O fitness de
uma dada caracterstica medido pelo nmero de descendentes de pessoas afectadas que
sobreviveram at idade reprodutiva em relao a uma populao controlo apropriada, por
outras palavras, traduz-se pela capacidade de um indivduo com um determinado fentipo de
doar genes gerao seguinte.
Incidncia e prevalncia de doenas genticas e fitness- pgina 2.
DIVERSIDADE E PATOLOGIA
1. Mecanismos e consequncias de mutao_____________________
Mecanismos de mutao e classificao de mutaes
C Cl la as ss si if fi ic ca a o o d de e m mu ut ta a e es s: :
Mutaes pontuais = quando envolve substituio, deleco ou insero de apenas um
nucletido.
Substituio de bases: Mais comuns: transio: substituies de uma pirimidina (C ou T) por
outra pirimidina ou de uma purina (A ou G) por outra purina; translocao: substituies de
uma purina por uma pirimidina e vice-versa.
Expanso de tripletos: Triplet expansion is caused by slippage during DNA replication. Due to
the repetitive nature of the DNA sequence in these regions, 'loop out' structures may form
Sebenta de Gentica
FCML
64
during DNA replication while maintaining complementary base paring between the parent
strand and daughter strand being synthesized. If the loop out structure is formed from
sequence on the daughter strand this will result in an increase in the number of repeats.
However if the loop out structure is formed on the parent strand a decrease in the number of
repeats occurs. It appears that expansion of these repeats is more common than reduction.
Generally the larger the expansion the more likely they are to cause disease or increase the
severity of disease. This property results in the characteristic of anticipation seen in
trinucleotide repeat disorders. Anticipation describes the tendency of age of onset to decrease
and severity of symptoms to increase through successive generations of an affected family
due to the expansion of these repeats. At present there are 14 documented trinucleotide
repeat disorders that affect humans (ver pg. 121).
Quadro 5: Classificao das mutaes
Conceito de pr-mutao (expanses instveis de tripletos): A heritable change, such as a
trinucleotide repeat expansion, that has no phenotypic effect but greatly increases the
likelihood of a further change at the altered site, resulting in a characteristic phenotype.
M Me ec ca an ni is sm mo os s d de e m mu ut ta a o o
O DNA pode ser danificado por agentes fsicos, qumicos ou ambientais e os danos
resultantes, podem ser classificados em 4 tipos:
Sebenta de Gentica
FCML
65
1. Alterao de 1 base- despurinao, desaminao, alquilao, insero ou deleco de
um nucletido.
2. Alterao de 2 bases- radiaes UV, que induzem a formao de dmeros de timina.
3. Quebra da cadeia- radiao ionizante.
4. "Cross-linkage"- ocorre entre bases, na mesma ou em cadeias opostas, entre o DNA e
molculas proteicas.
T Ta au ut to om me er ri ia a d de e b ba as se es s
Uma caracterstica importante do DNA a posio dos tomos de hidrognio nas bases de
purina e pirimidina que, apesar de poderem mudar de posio segundo movimentos
tautomricos, ocupam posies preferenciais. Os tomos de nitrognio ligados aos aneis de
purina e pirimidina esto normalmente sob a forma amino (NH
2
) e raramente assumem a
forma imino (NH). Os tomos de oxignio ligados ao carbono 6, na guanina e timina,
normalmente assumem a conformao ceto (C-O) e raramente, a conformao enol (COH).
essencial para a funo biolgica que os tomos de hidrognio do DNA ocupem posies
relativamente estveis, caso contrrio, a adenina poderia ligar-se citosina e a guanina
timina e a sequncia de bases das duas cadeias deixaria de ser obrigatoriamente
complementar. Nesse caso, o DNA no poderia funcionar como uma molcula gentica, uma
vez que a relao complementar entre as cadeias opostas, que lhe confere a capacidade
de replicao.
As bases podem apresentar-se, sob 2 formas distintas:
-Lactnicas- predomina a adenosina e a citosina.
-Lactnicas- predomina a timina e a guanina.
Por exemplo, a adenina pode alterar a sua forma tautomrica habitual, passando de lactnica
a lactnica e, em vez de emparelhar com a timina, emparelha com a citosina.
E Er rr ro os s d de e p pr ro oo of f- -r re ea ad di in ng g e e e em mp pa ar re el lh ha am me en nt to os s e er rr ra ad do os s
Erros na aco desta enzima tambm podem estar na base de algumas mutaes. Quando
os desoxirribonucletidos apropriados esto disponveis, as DNA-polimerases desempenham
a sua funo com a mxima fidelidade. Algumas mutaes ocorrem durante a replicao do
DNA (frequncia de cerda de 10
-8
), no entanto, estas alteraes esto limitadas pela grande
fidelidade evidenciada pela aco das DNA-polimerases e pelas propriedades de
exonuclease destas enzimas ou de outras auxiliadoras, que permitem a realizao das
funes de proofreading e de reparao. Quando h erros de proof-reading geralmente h
formao de expanses instveis dos tripletos.
Sebenta de Gentica
FCML
66
Deslizamentos da DNA polimerase em regies repetitivas
Ver expanso de tripletos pg. 64
D De es sa am mi in na a o o e es sp po on nt t n ne ea a d da a 5 5- -m me et ti il l c ci it to os si in na a e e h hi ip pe er rm mu ut ta ab bi il li id da ad de e r re eg gi i e es s C Cp pG G
In the human genome, an estimated 56% of genes are associated with CpG sequences They
include all examples of housekeeping genes and genes that are widely expressed, and a
significant portion, perhaps about 40% of genes which show tissue-specific or restricted
expression patterns. The dinucleotide CpG is a hotspot for mutation in the human genome
as a result of the modification of the 5' cytosine by cellular DNA methyltransferases and the
consequent high frequency of spontaneous deamination of 5-methyl cytosine (5 mC) to
thymidine. DNA methylation thus contributes significantly, albeit indirectly, to the incidence of
human genetic disease.
Fig. 30: The CpG dinucleotide is underrepresented in vertebrate DNA because it is prone to methylation and
deaminated 5-methylcytosine is subject to ineffective DNA repair (A) Cytosine occurring in the sequence 5-CpG-3
is a target for methylation at the 5 carbon atom. The deaminated products of cytosine and its methylated derivative
5-methylcytosine are differentially recognized by DNA repair enzymes. (B) Deamination of 5-methylcytosine in the
sequence CpG results in conversion of CpG dinucleotides to TpG and CpA dinucleotides. (C) The methylation
pattern of CpGs is perpetuated by a requirement for the specific methylase to recognize a hemimethylated target
sequence. The sequence CpG has dyad symmetry. Following methylation of a hemimethylated target (i.e.
methylated on one strand only), the two methylated strands will separate at DNA duplication and act as templates
for the synthesis of two unmethylated daughter strands. The resulting daughter duplexes will now provide new
hemimethylated targets for continuing the same pattern of methylation.
M Mu ut ta a e es s i in nd du uz zi id da as s p po or r m mu ut ta ag g n ne eo os s f f s si ic co os s e e q qu u m mi ic co os s
R Ra ad di ia a o o i io on ni iz za an nt te es s e e U UV V
Sebenta de Gentica
FCML
67
So mutagnios fsicos.
Radiaes ionizantes (Radiaes X e Radiaes ) Os mecanismos pelos quais actuam
levando mutao, foram mais bem estudados com base nas vtimas e seus descendentes
de Hiroshima e Nagasaki, bem como de acidentes com centrais nucleares (Chernobyl).
As radiaes ionizantes podem actuar:
- directamente na cadeia de DNA, induzindo a libertao de um electro de um tomo de uma
base e esta, assim ionizada, deixa de poder emparelhar normalmente, participando em
ligaes atpicas (ex.: a timina ionizada emparelha com a guanina).
- indirectamente, a radiao ionizante promove a radilise de H
2
O, o que leva formao de
radicais livres de O
2
(sobretudo OH
.
, o qual possui grande potencial oxidativo). Este radical
tem a capacidade de iniciar reaces em cadeia, actuando ao nvel das membranas
promovendo a peroxidao lipdica, com formao do composto malonil aldedo, que actua a
nvel nuclear, promovendo quebras no DNA. Desencadeia, sobretudo, quebras
cromossmicas induzindo fenmenos como a deleco, translocao, formao de
dicntricos e fragmentos acrocntricos. Os radicais livres podem tambm originar aductos de
DNA.
Alguns xenobiticos podem, ainda, actuar como haptenos, ou seja, como molculas que,
sozinhas no tm a capacidade de estimular a sntese de anticorpos, mas que, associadas a
molculas proteicas so capazes de se ligar queles, caso sejam formados. Esta situao
pode provocar leses celulares graves.
Radiaes UV A radiao UV induz a formao de dmeros pirimidina-pirimidina
(predominantemente T-T). Um dmero de timina forma-se quando 2 nucletidos de timina
sucessivos, de uma mesma cadeia, se ligam covalentemente por um ciclo de 4 carbonos
(ciclobutano). As duas timinas deixam de estar alinhadas correctamente ao longo da dupla
hlice e rompem-se as ligaes de hidrognio com as bases complementares, perdendo o
contacto com estas. Para alm disso, o esqueleto do DNA sofre uma sbita torso de cada
um dos lados do dmero de timina e 2 bases da mesma cadeia perdem tambm o contacto
com as suas bases complementares, da cadeia oposta. A radiao UV pode originar aductos
de DNA. Reparao por NER.
A Ag ge en nt te es s q qu u m mi ic co os s i in nt te er rc ca al la an nt te es s e e a an n l lo og go os s d de e b ba as se es s
Agentes intercalantes (ou anlogos a bases): os compostos orgnicos que possuam uma
estrutura aromtica de dimenses e estrutura apropriadas podem ser inseridos entre pares
de bases na dupla cadeia de DNA, fenmeno que se designa por intercalao. Durante este
processo, os pares de bases adjacentes so separados para permitir a insero do sistema
Sebenta de Gentica
FCML
68
intercalante, levando elongao da dupla hlice. Isto perturba a continuidade das
sequncias de bases do DNA e o modelo de transcrio para a DNA polimerase, levando, na
replicao seguinte, formao de uma molcula de DNA com uma base adicional (mutao
do tipo frameshift). Exemplos de agentes intercalantes so o brometo de etdio e acridinas.
A Ag ge en nt te es s q qu u m mi ic co os s a al lq qu ui il la an nt te es s e e a ar ri il la an nt te es s. . A Ad du uc ct to os s d de e D DN NA A
Agentes alquilantes: (substncias que podem doar grupos metilo ou etilo). Estes agentes
alteram a estrutura das bases, podendo igualmente quebrar ligaes fosfodister, levando
fragmentao do DNA. Para alm disto, outros agentes da mesma natureza podem interagir
covalentemente com ambas as cadeias, levando formao de pontes covalentes entre elas
formando aductos qumicos. Reparao por NER.
Agentes arilantes: podem ser endgenos formados naturalmente durante os processos
metablicos normais, tais como radicais livres de O
2
ou exgenos (como a alimentao e a
poluio). Os exgenos podem ainda ser divididos em: naturais (toxinas animais ou vegetais)
e industriais.
Teste de AMES: O teste de Ames tem sido o mais utilizado para avalio da eventual
mutagenicidade de compostos qumicos. Tal deve-se sua relativa facilidade de execuo,
baixos custos e a uma elevada sensibilidade relativamente a cancergenos conhecidos, se
bem que no seja sensvel a alguns tipos de agentes cancergenos tal como compostos
policlorinados e asbestos. Utiliza estirpes de bactrias Salmonella typhimurium
genticamente modificadas, contendo uma mutao no opero histidina (his -), que as torna
incapazes de sintetizar este aminocido. Nas estirpes utilizadas no teste de Ames, um
pequeno nmero de clulas - revertentes espontneos - tende a reverter a mutao inicial,
tornando-se assim capazes de crescer num meio contendo apenas uma fonte de carbono e
sais minerais. Para alm da mutao no opero responsvel pela sntese de histidina, as
estirpes utilizadas no teste de Ames contm outras mutaes (rfa e AuvrB) que levam a um
aumento da sua sensibilidade relativamente a mutagneos. Neste teste possvel estudar
duas variantes: o efeito directo do mutagnio, pela mutao directa do DNA; e o seu efeito
indirecto, pela aco dos seus metabolitos (que se formam a partir da degradao do
mutagnio no sistema microssomal, que neste caso, in vitro, corresponde a uma fraco
microssomal de fgado de rato adicionado preparao).
Metabolizao de pr-mutagneos (Metabolismo de fase 1 e de fase 2)
Existem produtos qumicos que, para exercerm a sua actividade lesiva sobre o material
gentico, necessitam de activao metablica prvia. Considera-se, ento, que estes
Sebenta de Gentica
FCML
69
produtos actuam indirectamente sob a forma de pr-mutagnios (pro-carcinognios),
geralmente terobiticos (substncias que estimulam o metabolismo do organismo e que
necessitam de activao metablica). A maior parte destes compostos apresentam um
acentuado carcter lipoflico, travessando facilmente as membranas biolgicas. Nos
organismos superiores estes compostos no podem ser excretados seno aps serem
transformados em compostos mais polares. Este tipo de reaces predominatentemente
catalisado pelo sistema de mooxigenases dependentes do citocromo P450. De facto, a
expresso citocromo P450 designa uma superfamlia de protenas que catalisa um elevado
nmero de reaces, em que se incluem reaces envolvidas na biossntese de cidos
biliares e hormonas, o metabolismo de certos cidos gordos e de prostaglandinas e ainda a
oxidao de um grande nmero de compostos exgenos. O metabolismo destes compostos
pode envolver um elevado nmero de reaces, das quais podem resultar, quer compostos
capazes de reagir com centros nucleoflicos de biomolculas, quer compostos no
genotxicos. Assim, considera-se a existncia de duas fases distintas no metabolismo dos
pr-genotxicos: uma primeira fase em que intervm um conjunto de enzimas designadas por
enzimas de fase I (sistema de mooxigenases dependentes do citocromo P450, reductases e
hidrolases), que leva activao do genotxico (denominado nesta dase metabolito)
aumentando o seu carcter electroflico de modo a facilitar a sua excreo; e uma segunda
fase em que intervm um grupo de enzimas designadas por enzimas de fase II
(transferases), que conduz conjugao dos metabolitos formados na fase I com outros
compostos de acentuado carcter hidroflico formando molculas hodrossolveis, com vista a
facilitar a sua excreo. Isto acontece porque os xenobiticos so muitas vezes compostos
apolares e, por isso, no podem ser facilmente excretados a nvel renal por no serem
hidrossolveis. Deste modo, tenderiam a acumular-se indefinidamente em tecidos com
grande densidade de compostos igualmente apolares, como tecido adiposo. Aps sofrerem
metabolizao, os compostos tornam-se, ento, polares e mais solveis, passveis de ser
excretados na blis, embora se possam, no processo, tornar carcinognicos e actuar a nvel
nuclear, lesando o DNA.
O Fgado o rgo mais importantes na biotransformao de xenobiticos.
Sebenta de Gentica
FCML
70
Fig. 31: Biotransformao
Papel da biotransformao na etiologia do cancro:
Sebenta de Gentica
FCML
71
A A s su up pe er r f fa am m l li ia a g g n ni ic ca a d do os s c ci it to oc cr ro om mo os s P P4 45 50 0 ( (g ge en ne es s C CY YP P) )
Os seres vivos esto continuamente expostos a compostos sem qualquer valor nutritivo. O
modo como este tipo de compostos pode ser metabolizado varia entre os diferentes tipos de
seres vivos. Enquanto que algumas bactrias so capazes de convereter estes compostos
em metabolitos utilizveis, os mamferos evoluram no sentido de adquirirem mecanismos
para a sua eliminao. A maior parte destes compostos apresentam um acentuado carcter
lipoflico, travessando facilmente as membranas biolgicas. Nos organismos superiores estes
compostos no podem ser excretados seno aps serem transformados em compostos mais
polares. Este tipo de reaces predominatentemente catalisado pelo sistema de
mooxigenases dependentes do citocromo P450. De facto, a expresso citocromo P450
designa uma superfamlia de protenas que catalisa um elevado nmero de reaces, em que
se incluem reaces envolvidas na biossntese de cidos biliares e hormonas, o metabolismo
de certos cidos gordos e de prostaglandinas e ainda a oxidao de um grande nmero de
compostos exgenos. Da superfamlia dos citocromos P450 so conhecidas, at data pelo
menos 36 famlias, 12 das quais presentes em mamferos. A designao dos diferentes
genes desta superfamlia foi proposta por Nerbert (1991), tendo como base a sua evoluo
biolgica. Assim, para a designao do gene, utiliza-se o prefixo CYP denotando citocromo
P450, seguindo-se um nmero rabe denotando a famlia, uma letra indicando a subfamlia e,
finalmente um segundo nmero rabe para indicar o gene individual. O sistema das
mooxigenases dependentes do citocromo P450 est localizado na membrana do retculo
endoplasmtico, num complexo multienzimtico que inclui igualmente uma redutase. Em
mamferos, este complexo multienzimtico est presente em vrios tecidos, como o pulmo e
Sebenta de Gentica
FCML
72
o intestino delgado, encontrando-se em maior quantidade no fgado. Apesar da grande
diversidade de reaces catalisadas pelo citocromo P450, um grande nmero resulta na
insero de um tomo de oxignio entre uma ligao R-H, conforme indicado no esquema
seguinte: R-H + O
2
+ NADPH + H
+
ROH + NADP
+
+ H
2
O.
O complexo multienzimtico recebe os electres do NADPH ou do NADH, que passam
atravs de uma flavoprotena que catalisa uma reaco de oxirreduo, atravs do citocromo
b5 ou directamente para o prprio citocromo P450. Este, em combinao com o oxignio e
um dos numerosos substratos, forma um complexo trimolecular, que resulta na oxigenao
do substrato. Um elevado nmero de genotxicos manifesta apenas essa actividade aps
sofrer metabolizao por esta via, sendo, por este motivo, designados por pr-genotxicos. O
metabolismo destes compostos pode envolver um elevado nmero de reaces, das quais
podem resultar, quer compostos capazes de reagir com centros nucleoflicos de
biomolculas, quer compostos no genotxicos (metabolismo fase I e II). Vrios factores
podem alterar o metabolismo dos xenobiticos, qualitativa e quantitativamente, uma vez que
os diferentes genes do citocromo P450 so indutveis por compostos exgenos. Um dos
determinantes da actividade do citocromo P450 parece ser a alimentao, j que as
diferentes actividades deste sistema enzimtico so moduladas por diferentes factores
nutricionais, entre os quais se citam os teores em protenas, lpidos, hidratos de carbono e
micronutrientes. No entanto, outros compostos so igualmente capazes de induzir o sistema
de monooxigenases dependentes do citocromo P450, entre os quais se encontram o TCDD
(2,3,7,8-tetraclorodibenzeno-p-dioxina), o Aroclor 1254 (uma mistura de pentaclorobifenil com
tetra, hexa e heptaclorobifenil), o 3-metil-colantreno e o fenobarbital. O citocromo P450 pode
ainda catalisar a reduo univalente do substrato, gerando-se um radical centrado em tomos
de carbono, oxignio, azoto e enxofre. Os compostos assim formados podem reagir
directamente com biomolculas ou ainda com o oxignio, levando, neste caso, formao do
radical superxido. Para alm do fgado, outros orgos, ainda que em menor extenso,
contm enzimas capazes de catalisar reaces envolvidas na activao de pr-genotxicos.
Em clulas do epitlio do tubo digestivo foram identificadas diversas enzimas envolvidas na
activao de xenobiticos. A flora intestinal humana compreende cerca de quatrocentas
espcies bacterianas, responsveis por 30% da massa fecal. A composio relativa deste
verdadeiro sistema ecolgico dependente de diversos factores, tais como a idade e a
alimentao. Algumas destas espcies bacterianas so capazes de catalisar um grande
nmero de reaces envolvidas na activao metablica de xenobiticos previamente
metabolizados no fgado, conjugados com molculas polares, no activos face ao DNA, ao
serem excretados para o intestino atravs da blis podem ser hidrolisados por aco das
glucuronidases bacterianas, podendo levar formao de genotxicos directos. Para alm
Sebenta de Gentica
FCML
73
disso, algumas bactrias intestinais contm actividades capazes de hidrolisar genotxicos
naturais que se encontrem na forma de glicsidos. Assim, compostos genotxicos, como a
cicasina e glicsidos de flavonides, cuja actividade genotxica apenas se manifesta aps
hidrlise do glicsido, podem ser activados directamente no tubo digestivo, originando
agliconas mutagnicas. Vrias espcies do gnero Bacteroides, presentes na flora intestinal
humana so capazes de metabolizar plasmalognios (uma classe de lpidos) em
fecapentenos. Estes compostos possuem reconhecida aco genotxica e carcinognica e
tm sido associados mutagenicidade observada em fezes de diferentes populaes. Os
mecanismos de leso gentica por estes compostos no esto ainda esclarecidos, existindo
vrios dados que apontam para o envolvimento de epcies activas de oxignio na sua
genotoxicidade.
Inducibilidade e susceptibilidade a cancro- ver gentica do cancro
Significado e consequncias de mutaes
E Em m r re eg gi i e es s e ex xt tr ra a- -g g n ni ic ca as s: : a an n n ni im ma as s ( (S SN NP Ps s, , V VN NT TR R e e S ST TR R) )
SNP: Single Nucleotide Polymorphism is a small genetic change, or variation, that can occur
within a person's DNA sequence. SNP variation occurs when a single nucleotide, such as an
A, replaces one of the other three nucleotide lettersC, G, or T. An example of a SNP is the
alteration of the DNA segment AAGGTTA to ATGGTTA, where the second "A" in the first
snippet is replaced with a "T". On average, SNPs occur in the human population more than 1
percent of the time. Because only about 3 to 5 percent of a person's DNA sequence codes for
the production of proteins, most SNPs are found outside of "coding sequences". SNPs found
within a coding sequence are of particular interest to researchers because they are more likely
to alter the biological function of a protein. For geneticists, SNPs act as markers to locate
genes in DNA sequences. Say that a spelling change in a gene increases the risk of suffering
from high blood pressure, but researchers do not know where in our chromosomes that gene
is located. They could compare the SNPs in people who have high blood pressure with the
SNPs of people who do not. If a particular SNP is more common among people with
hypertension, that SNP could be used as a pointer to locate and identify the gene involved in
the disease.
Os SNPs incluem os RFLPs mas tambm outros que no alteram stios de restrio. Embora
menos informativos que os microssatlites, os SNPs so mais adequados nas contagens
automatizadas de grande escala.
Sebenta de Gentica
FCML
74
VNTR: Variable Number Tandem Repeat so microssatlites, minissatlites e megassatlites,
ou seja, arranjos de sequncias repetidos em tandem, que frequentemente variam entre as
pessoas quanto ao nmero de unidades repetidas. O termo VNTR em geral usado com o
significado especfico de minissatlites (ver pg. 8).
STR: Short Tandem Repeat in DNA occurs when a pattern of two or more nucleotides are
repeated and the repeated sequences are directly adjacent to each other. The pattern can
range in length from 2 to 16 base pairs (bp) (for example (CATG)
n
in a genomic region) and is
typically in the non-coding intron region. By identifying repeats of a specific sequences, it is
possible to create a genetic profile of an individual. There are currently over 10,000 published
STR sequences in the human genome. STR analysis has become the prevalent analysis
method for determining genetic profiles in forensic cases.
E Em m r re eg gi i e es s c co od di if fi ic ca an nt te es s: :
Mutaes silenciosas a leitura do cdigo gentico degenerado, e mesmo que ocorra uma
mutao em determinado nucleotido, pode no ocorrer nenhuma alterao na traduo da
protena.
Fig. 33: Mutao silenciosa
Mutaes Missense alterao de um aa. Podem ser perigosas (aa de carga e/ou estrutura
diferente) ou neutras (degenerescncia paralela do cdigo).
Fig. 34: Mutao missense
Mutaes Nonsense introduo precria de um codo STOP. Alterao de um aminocido
para um codo STOP, que leva produo de uma protena truncada. Se o novo codo
Sebenta de Gentica
FCML
75
STOP estiver prximo do fim, a protena no tem o comprimento que deveria mas a
mutao quase imperceptvel.
Fig. 35: Mutao Nonsense
non-sense mediated RNA decay reconhece o STOP mal localizado, onde se agrega uma
srie de protenas e levam a um decay e a um recycling das protenas. O reconhecimento por
estas protenas pode ocorrer no ncleo.
Mutaes Frameshift alterao do cdigo de leitura: passa a ler de 3 em 3, formando
codes diferentes. Normalmente produz uma protena truncada. Ex.: doena recessiva
fibrose cstica.
Fig. 36: Mutao Frameshift
Splicing mutation- splincing alternativo que deixa de funcionar; destruio ou criao de um
local de splice; traduo de exes ou activao de locais de cryptic splice.
Fig. 37: Splicing mutation
E Em m r re eg gi i e es s r re eg gu ul la ad do or ra as s
As mutaes em regies reguladoras, no codificantes, podem alterar a taxa de transcrio
do gene e originar patologia por alterao da quantidade de protena produzida. Isto acontece
Sebenta de Gentica
FCML
76
se para a funo da protena a quantidade for um factor determinante. o caso das
talassmias, doenas provocadas por um desequilbrio das quantidades das cadeias e da
globina. A fig. 38 representa o gene da -globina, a localizao das vrias mutaes
associadas -talassmia e respectivos mecanismos patognicos. As mutaes -29AG e -
88CT localizam-se na regio reguladora do gene da -globina e causam -talassmia.
Alteraes epigenticas da regio reguladora, como a metilao, tambm inibem a
transcrio e so um mecanismo frequente de inactivao de genes supressores tumorais no
cancro.
Consequncias patolgicas em cada tipo de mutao
Os efeitos das mutaes vo depender das alteraes que induzem na funo da protena e
da funo da protena ao nvel da clula ou do organismo.
Fig. 39: Efeito das mutaes
Fig. 38: gene da -globina, a localizao das
vrias mutaes associadas -talassmia e
respectivos mecanismos patognicos
Sebenta de Gentica
FCML
77
As mutaes nonsense podem ter diferentes consequncias: 1- originarem um mRNA
instvel, degradado pelo sistema NMD; 2 originarem um polipeptdeo truncado, que poder
ou no ter algum grau de funo ou que possa interferir com polipeptdeos normais (caso das
protenas multimricas e da dominncia negativa).
As mutaes tipo frameshift frequentemente tm efeitos semelhantes s nonsense porque a
alterao da grelha de leitura que introduzem acaba por criar codes stop prematuros.
As mutaes localizadas em sequncias envolvidas no splicing podem originar reteno de
intres se o site receptor (SR) de splicing no for reconhecido e o intro no for excisado ou
eliminao de exes se o spliceosoma juntar sites dadores (SD) e receptores no contguos.
Frequentemente associam-se-lhes o efeito de frameshift e formao de codes stop
prematuros a jusante.
Mtodos de deteco de Mutaes:
Um estudo molecular (PCR, southern blotting, sequenciao) identifica uma mutao pontual
no genoma.
SSCP- mtodo de electroforese que consegue dstinguir cadeias de DNA, diferindo em
apenas um nucletido. As cadeias simples so sujeitas electroforese e a diferente
conformao espacial da cadeia mutada em relao cadeia nativa permite-lhe migrar
diferencialmente no gel.
RFLPs (restriction fragment lenght polimorphisms)- as enzimas de restrio tm sequncias
de reconhecimento especficas no DNA. Alteraes da sequncia do DNA genmico que
ocorrem por mutao hereditria ou nova, promovem a criao ou a abolio de locais de
clivagem, alterando o tamanho de um ou mais fragmentos de DNA que se evidencia no
Southern blotting e hibridao com uma sonda de DNA clonada.
VNTRs (variable number of tandem repeats)- um tipo de polimorfismo criado por um
arranjo repetitivo de mltiplas cpias de pequenas sequncias de DNA. Consistem numa
srie de fragmentos allicos, relacionados entre si por um n varivel de sequncias repetidas
em tandem, no intervalo de dois locais de restrio. Esta classe de RFLPs foi denominada
hipervarivel, pois tais loci caracterizam-se por muitos alelos. So utilizados como
marcadores em estudos de linkage e em DNA fingerprinting para testes de paternidade e
medicina forense. As variaes nos locais de restrio do DNA verificados desta forma,
denominam-se RFLPs.
Sebenta de Gentica
FCML
78
Tipos de mutao mais frequentes em patologia humana
Fig. 40: mutaes mais frequentes
Impacto em patologia gentica da taxa de neo-mutao humana
A taxa de neomutao, estimada pela frequncia de doenas genticas verificadas sem se
deverem a transmisso familiar relativamente elevada na espcie humana, sendo de cerca
de 80%.
2. Mecanismos protectores da fidelidade informativa______________
Sistemas enzimticos de reparao do DNA
R Re ep pa ar ra a o o p pr r - -r re ep pl li ic ca at ti iv va a p pe el la as s D DN NA A- -p po ol li im me er ra as se es s
Funo de proofreading da DNA polimerase - A DNA polimerase tem uma funo de
proofreading. Com efeito, examina o resultado de cada polimerizao que catalisa antes de
prosseguir com a prxima. Se houver erro de emparelhamento activada a enzima 3'5'
exonuclease do complexo DNA polimerase, que cliva o ltimo nucletido da cadeia e o
substitui.
R Re ep pa ar ra a o o p p s s- -r re ep pl li ic ca at ti iv va a p po or r r re ec co om mb bi in na a o o ( (R Re ec co om mb bi in na at ti io on na al l R Re ep pa ai ir rs s) )
necessria para corrigir quebras nas fitas duplas. O mecanismo usual um processo
semelhante converso gnica (reparao por recombinao), em que uma fita simples de
um cromossoma homlogo invade a hlice de DNA danificada. Alternativamente, as
extremidades quebradas so religadas sem ter em considerao as suas sequncias, uma
medida desesperada, que passvel de causar mutaes. A maquinaria eucaritica para
Sebenta de Gentica
FCML
79
reparao por recombinao bem menos definida do que os sistemas de reparao por
exicso. Os genes humanos envolvidos nessa rota incluem o NBS (mutado na sndrome de
Nimegen), BLM (mutado na sndrome de Bloom) e os genes de susceptibilidade ao cncro da
mama, BRCA2 e possivelmente o BRCA1.
Reparao por exciso; Glicosilases e endonucleases de locais apurnicos; NER,
BER, MMR e NHEJ
NER (Nucleotide Excision Repair): remove dmeros de timna e aductos. um processo
sequencial que consiste, em termos genricos, em quatro passos consecutivos: inciso,
exciso, ressntese e ligao. O primeiro passo o de reconhecimento da alterao
conformacional do DNA (distoro) por um sistema de endonucleases constitudo, nas
clulas eucariticas por 16-17 polipptidos, sendo o processo especfico para cada tipo de
leso presente. tambm a etapa limitante de todo o processo. A exciso consiste na
remoo do fragmento de DNA lesado (clivagem fosfodiestersica nas extremidades da
leso) pelo mesmo complexo enzimtico, deixando uma lacuna na cadeia de DNA. A
designao formal dos polipptidos constituintes deste complexo pode ser ERCC (excision
repair component) ou XP (xeroderma pigmentosum), devendo-se esta ltima designao ao
facto de as protenas XPA a XPG terem sido identificadas como constituintes de sete grupos
de complementao de pacientes com xeroderma pigmentosum. O processo de exciso de
nucletidos inicia-se com o reconhecimento do local lesado por parte da protena XPA,
seguido do recrutamento pela mesma dos factores XPF e ERCC1, que dimerizam e se ligam
a ela. Seguidamente, a protena de replicao HSSB (RPA) liga-se simultaneamente ao
dmero XPF-ERCC1 e ao DNA lesado. Um aspecto surpreendente deste tipo de reparao de
DNA que a estruturao do complexo de endonucleases requer a presena de um
complexo enzimtico adicional constitudo por oito polipptidos e conhecido por factor geral
de transcrio TFIIH. Com efeito, este, para alm de ser um factor essencial para a iniciao
da transcrio -o tambm para a reparao, pois constitudo pelas helicases XPB e XPD,
que, evidentemente, no actuam somente na reparao, mas tambm catalisam o
desenrolamento do DNA para a transcrio. Esta ntima associao sugere que os processos
de reparao e de transcrio no so totalmente distintos, podendo estar estreitamente
relacionados, pelo que isto daria clula uma via atravs da qual poderia regular
estreitamente os dois processos. Seguidamente, TFIIH interage com XPC e o complexo
recrutado para o local da leso pela XPA, juntando-se, posteriormente ainda a endonuclease
XPG. Esta ltima completa o sistema exonuclesico, sendo responsvel pela clivagem na
extremidade 3 do oligopptido a excisar (3 resduos de nucletidos a jusante da leso). Por
sua vez, a clivagem da extremidade 5 da responsabilidade do dmero XPF-ERCC1,
Sebenta de Gentica
FCML
80
normalmente o 25 resduo a montante. Seguidamente, uma protena associada polimerase
o, a PCNA (proliferating cell nuclear antigen) liberta todas as subunidades do complexo, bem
como o oligonucletido removido, ocorrendo, posteriormente a ressntese da lacuna formada,
pelas polimerases o e c e o factor de replicao RFC.
Fig. 41: Anlise do processo da figura:
1. Etapa de reconhecimento do DNA danificado pela XPA (liga-se directamente ao DNA ou atravs da
protena RPA)
2. Desenrolamento atravs da helicase formando uma bolha no DNA de +- 30 nucletidos. Funo do
factor geral de transcrio TFII do qual fazem parte a XPB e a XPD
3. Exciso pela nuclease que retira grandes pores de DNA (ex. dmeros de timina e aductos). O ECRR +
(XPF na extremidade 5 e XPG na extremidade 3) quebram as ligaes entre a poro danificada e no
danificada do DNA
4. DNA polimerase: acrescenta bases
5. DNA ligase: fecha o corte entre novo nucletido e cadeia
BER (Base Excision Repair) e glicosilases e endonucleases de locais apurnicos:
Remove bases anormais. A despurinizao do DNA, que ocorre espontaneamente a 37C
devido labilidade trmica da ligao N-glicosdica das purinas muito frequente, ocorrendo
em cada dia cerca de 5000 a 10,000/clula. Enzimas especficas tm a capacidade de
reconhecer estes locais apurnicos e introduzir directamente a purina adequada que falta,
sem interromper a ligao fosfodister. Para alm desta alterao, a citosina, a adenina e a
guanina so igualmente susceptveis de sofrer desaminao espontnea, transformando-se,
respectivamente, em uracilo, hipoxantina e xantina. No entanto, devido maior estabilidade
da ligao N-glicosdica nas pirimidinas do que nas purinas, a despirimidizao no to
frequente nestas ltimas. Por outro lado, vrios mutagnios qumicos e radiaes ionizantes
podem provocar alteraes profundas nas bases do DNA. Estas bases atpicas so
reconhecidas e hidroliticamente removidas por enzimas designadas por DNA glicosilases.
Esta remoo serve de marcador do local da leso, que reconhecido por endonucleases de
lucais apurnicos (ou apirimidnicos), que clivam a ligao fosfodister correspondente da ose
absica. Seguidamente, a aco combinada da DNA polimerase I e da DNA ligase leva
Sebenta de Gentica
FCML
81
restaurao da cadeia danificada. Quando a reparao por exciso no desempenha
correctamente o seu papel, a forquilha de replicao corre o risco de ficar bloqueada, se
certos mecanismos de reforo no existirem fisiologicamente.
Fig. 42: Anlise do processo da figura:
1. Glicosilase: retira a base, ficando uma zona sem base.
2. Endonuclease: corta ligao ose-fosfato na posio da base retirada
3. DNA polimerase: acrescenta base
4. DNA ligase III: fecha o corte entre novo nucletido e cadeia
MMR (MisMatch Repair): na reparao do DNA essencial diferenciar as regies normais
das regies aberrantes. Esta informao encontra-se, por vezes, no prprio defeito (se a
base est modificada, ex: dmero de timina). No entanto, se o erro no afectar a estrutura
conformacional da dupla hlice, a informao errnea identificada atravs dos grupos
metilo que se encontram nos resduos de adenina e nas sequncias de GATC da cadeia
matriz. Com efeito, aps a replicao, s na cadeia matriz que o DNA est metilado em
muitos locais, pois a nova cadeia de DNA ainda no teve tempo para tal, j que a metilao
um processo demorado. Logo, a enzima de reparao corta as bandas no metiladas para
remover o erro nucleotdico, deixando o DNA padro intacto para que possa servir de modelo
de correco- mtodo utilizado na converso gnica (pgina 24). Os genes MSH2 e MSH6
so heterodmeros que reconhecem zonas onde h bases mal emparelhadas. O seu defeito
causa cancro do clon no poliposo hereditrio (HNPCC).
NHEJ (Non-homologous end joining) is a pathway that repairs double-strand breaks in DNA.
NHEJ is referred to as "non-homologous" because the break ends are directly ligated without
Sebenta de Gentica
FCML
82
the need for a homologous template. The NHEJ pathway is initiated in response to the
formation of a DNA double-strand break (DSB) induced by a DNA-damaging agent such as
ionizing radiation. First, the Ku70/80 heterodimer binds to the ends of the DSB. The catalytic
subunit of the DNA-dependent protein kinase (DNA-PKcs) is then recruited to DNA-bound Ku
to form the DNA-PK holoenzyme. The ends of the break are brought together as two
molecules of DNA-PK (one at each end of the break) are joined in a synaptic complex. Other
factors, such as polynucleotide kinase (PNK), Artemis, the MRE complex, hTdp1 or the
Werner Syndrome protein (WRN) may be required for processing of the DNA ends prior to
end rejoining, but exactly when processing takes place is not known. Following the formation
of the synaptic complex, the XRCC4/DNA ligase IV complex is recruited. Prior to end
rejoining, protein factors must be removed from the DNA. This may involve DNA-PK
autophosphorylation. After removal of the repair factors, the DNA ends are ligated and the
DNA is repaired. Both Mg-ATP and the protein kinase activity of DNA-PKcs are required for
NHEJ, probably through phosphorylation of DNA-PKcs and/or other proteins. In addition,
inositol hexaphosphate (IP6) stimulates end joining in vitro and binds to Ku, but its precise role
in NHEJ is unknown.
Fig. 43: Non-homologous end joining
Sebenta de Gentica
FCML
83
Sndromes de dfice de reparao e fragilidade cromossmica
A An ne em mi ia a d de e F Fa an nc co on ni i
Doena autossmica recessiva cujo gene do respectivo grupo de complementao A
(gene FAA) foi mapeado no brao longo do cromossoma 16. A principal causa da
anemia de fanconi a mutao do gene FAA. No entanto, h ainda mais 10 grupos
de complementao: C (cromossoma 9), D1 (crom. 3), D2, E (crom.6), F(11), G, I, J,
L, M. FANCB localiza-se no cromossoma X. Due to the similarities in the phenotypes
of the different FA complementation groups, it was reasonable to assume that all
affected genes interacted in a common pathway. Studies have shown that eight of
these proteins, FANCA, -B, -C, -E, -F, -G, -L and M assemble to form a core protein
complex in the nucleus. This complex has also been suggested to exist in cytoplasm
and its translocation into the nucleus is dependent on the nuclear localization signals
on FANCA and FANCE. Assembly is thought to be activated by replicative stress,
particularly that resulting from DNA damage caused by cross-linking agents (mitomycin
C or cisplatin) or reactive oxygen species (ROS). Indeed, FANCA and FANCG have
been observed to multimerize when a cell is faced with oxidative stress-induced
damage. Following assembly, the protein core complex activates FANCL protein
which acts as an E3 ubiquitin-ligase and monoubiquitinates FANCD2.
Monoubiquitinated FANCD2, also known as FANCD2-L, then goes on to interact with
a BRCA1/BRCA2 complex. Details of this interaction have yet to be discovered.
However, it is already known that similar complexes are involved in genome
surveillance and associated with a variety of proteins implicated in DNA repair and
chromosomal stability.
[11][12]
With a crippling mutation in any FA protein in the complex,
DNA repair has been shown to be much less effective, as can be seen from the
damage caused by cross-linking agents such as cisplatin, diepoxybutane and
Mitomycin C. It follows that tissues, as is the case in bone marrow, in which successful
cell replication is vital, will be severely affected by FA protein dysfunction where FA
leads to decreased haemopoiesis and bone marrow failure due to progenitor and stem
cell senescence. In another pathway responding to ionizing radiation, FANCD2 is
thought to be phosphorylated by protein complex ATM/ATR activated by double-
strand DNA breaks, and takes part in S-phase checkpoint control. This pathway was
proven by the presence of radioresistant DNA synthesis, the hallmark of a defect in the
S phase checkpoint, in patients with FA-D1 or FA-D2. Such a defect readily leads to
Sebenta de Gentica
FCML
84
uncontrollable replication of cells and might also explain the increase frequency of
AML in these patients. Corresponde a uma panmielopatia que se manifesta durante a
infncia, conduzindo a uma pancitopnia. Possuem, ainda, elevados nveis de HbF.
caracterizada por pancitopnia, atraso no crescimento, pigmentao acastanhada da
pele, falncia da medula ssea no final da infncia, embora apresente uma grande
heterogeneidade clnica e gentica.
A At ta ax xi ia a- -t te el la an ng gi ie ec ct ta as si ia a
Caracteriza-se por uma progressiva ataxia cerebelosa, reconhecida por volta dos 12 a 14
meses e telangiectasia oculo-cutnea (dilatao dos vasos subcutneos). O doente fica
confinado a uma cadeira de rodas antes da adolescncia. caracterstica a presena de uma
translocao no brao longo do cromossoma 14. As clulas apresentam menor capacidade
de reparao por exciso do DNA danificado (particularmente sensveis s radiaes
ionizantes), com consequente aumento da instabilidade cromossmica. Observa-se um
nmero elevado de quebras, gaps e dicntricos nas clulas destes indivduos aps a
exposio a radiaes de alta energia. Clinicamente h uma diminuio ou ausncia dos
nveis de IgA e IgG (muito teis para o diagnstico) e, portanto, diminuio das defesas a
agentes lesivos. Os doentes apresentam uma elevada tendncia para desenvolver leucemias
ou linfomas. O gene mutado parece ser o que codifica para a protena fosfatidilinositol-3-
quinase, uma das protenas implicadas na transduo de sinais mitognicos.
S S n nd dr ro om me e d de e L Ly yn nc ch h e e d d f fi ic ce e d de e r re ep pa ar ra a o o d de e e em mp pa ar re el lh ha am me en nt to os s e er rr r n ne eo os s
Ver pgina 173
X Xe er ro od de er rm ma a p pi ig gm me en nt to os su um m- - d d f fi ic ce e d de e N NE ER R
No Xeroderma Pigmentosum, doena gentica autossmica recessiva, os indivduos
afectados apresentam uma pele muito fina, frgil, envelhecida e enrugada prematuramente.
Apresentam fotodermatose induzida pela exposio a radiaes solares, caracterizadas por
reaces cutneas severas. Algumas formas so acompanhadas tambm de alteraes do
foro neurolgico. Os cancros cutneos aparecem em simultneo em vrios locais. A
exposio solar agrava as leses, precipitando a transformao maligna. Os fibroblastos
destes doentes, quando cultivados, revelam-se mais sensveis aos UV do que os fibroblastos
de indivduos normais. Para uma dose de radiao idntica, os fibroblastos dos doentes
apresentam muito mais dmeros de timina do que os dos indivduos normais. As clulas
destes indivduos tm menor capacidade de reparao do DNA danificado por radiao U.V.
(ou agentes qumicos), que se deve deficincia de endonucleases. As clulas so, assim,
Sebenta de Gentica
FCML
85
incapazes de reconhecer e cindir os locais afectados. Vrias alteraes enzimticas podem
levar a fentipos Xeroderma pigmentosum-like - heterogeneidade gnica. Com efeito foi
possvel determinar a existncia de sete grupos de complementao distintos, no qual se
inserem cerca de 80% dos indivduos com esta condio, o que levou descoberta de outras
tantas enzimas envolvidas no mecanismo de raparao por exciso, a que se deu o nome de
XPx, em que x representa uma letra de A a G.
S S n nd dr ro om me e d de e C Co oc ck ka ay yn ne e- - d d f fi ic ce e d de e N NE ER R
A sndroma de Cockayne uma doena autossmica recessiva associada a instabilidade
cromossmica. Nesta sndroma h reparao preferencial dos pontos de ancoragem do DNA
ao invlucro nuclear em vez das ansas (que se expressam mais e que so, normalmente,
reparadas primeiro) e que, portanto, acumulam erros, por outras palavras, h um defeito na
copulao da transcrio e da reparao, que providenciado pelo TFIIH e pela RNA
polimerase II. Observa-se uma incapacidade de reparao de dmeros de timina aps a
irradiao pelas radiaes UV e reduo da sntese de RNA aps essa leso. Deste modo,
so despoletados mecanismos que levam paragem do ciclo celular. Fenotipicamente h
progeria (senilidade precoce). um dfice de reparao no global (existem acumulaes de
leses no genoma que no so reparadas). Traduz-se por hipersensibilidade UV, o que pode
levar leso pigmentar da retina, envelhecimento precoce. As repetidas leses do endotlio
so factores que promovem o aparecimento de aterosclerose, com consequente hipertenso,
insuficincia renal e morte prematura. Apresentam ainda numerosas alteraes nerolgicas e
do desenvolvimento, designadamente, leucodistrofia, com desmielinizao e consequente
atraso mental. Por outro lado, observa-se morte neuronal, levando a demncia, alteraes da
marcha e incontinncia. As alteraes do desenvolvimento traduzem-se por nanismo, com
alterao das epfises de algumas falanges, membros desproporcionalmente longos,
desequilbrio muscular, etc. No apresenta tanta predisposio para o aparecimento de
melanomas. Salienta-se a presena de microcefalia, atraso no crescimento, atraso mental,
rash cutneo. Apresenta dois grupos complementares- dois genes mutados (ERCC8 e
ERCC6).
Fig. Genes envolvidos na sndrome de Cockayne
Sebenta de Gentica
FCML
86
T Tr ri ic co o- -t ti io od di is st tr ro of fi ia a- - d d f fi ic ce e N NE ER R
Indivduos com tricotiodistrofia apresentam deficincia de enxofre nos cabelos e nas unhas, o
que resulta no seu estado quebradio. Alm disso, esses pacientes apresentam atraso no
crescimento e atraso mental, alm de fotossensibilidade cutnea, embora sem predisposio
ao cncer. Determinados tipos de mutao de dois genes distintos: ERCC2/XPD e
ERCC3/XP podem causar a tricotiodistrofia. Esta sndrome pode tambm ser causada por
mutaes noutro gene, denominado TTDA e localizado em 6p25.3. Apresenta 3 grupos
complementares 3 genes mutados.
S S n nd dr ro om me e d de e B Bl lo oo om m e e o o d d f fi ic ce e d de e D DN NA A- -l li ig ga as se e
uma doena autossmica recessiva, com elevada incidncia nos judeus Ashakenazi. O
gene responsvel por esta sndroma o gene BLM, membro da famlia RecQ, o que leva
destabilizao de enzimas implicadas na sntese e na reparao do DNA, bem como
alteraes especficas da DNA ligase e da polimerase |. Estes aspectos levam a uma
hipermutabilidade, com diminuio da resposta da p53 exposio a mutagnios, o que leva
a uma hepersensibilidade a radiaes e ao aumento da incidncia de neoplasias.
Caracteriza-se pelo aparecimento de cromossomas tetrarradiais simtricos (resultante de
troca de cromtides entre cromossomas homlogos) por aco de agentes clastognicos,
elevada frequncia de troca de fragmentos entre cromtides irms (SCE), que cerca de 10
vezes superior aos pacientes com anemia de Fanconi. Resulta da deficincia em DNA-ligase
I e portanto, no ocorrncia da ligao entre os fragmentos de Okazaki durante a replicao e
dos oligonucletidos introduzidos durante a reparao. Os doentes apresentam atraso grave
no crescimento, baixo peso nascena, hipersensibilidade da pele a radiaes solares, leso
facial em forma de borboleta com telangiectasia (rash facial) e sensibilidade aumentada
infeces bacterianas devido a uma imunodeficincia. Tm elevada predisposio para o
cancro da pele, doena pulmunar obstrutiva crnica e diabetes.
DA DIVERSIDADE PATOLOGIA- EXPRESSO CLNICA E
MECANISMOS DE DOENA GENTICA
1. Patologia Gentica monognica______________________________
Fibrose qustica
Defects in CFTR protein are the cause of cystic fibrosis (CF); also known as mucoviscidosis.
CF is the most common genetic disease in the Caucasian population, with a prevalence of
Sebenta de Gentica
FCML
87
about 1 in 2'000 live births. Inheritance is autosomal recessive. CF is a common generalized
disorder of exocrine gland function which impairs clearance of secretions in a variety of
organs. It is characterized by the triad of chronic bronchopulmonary disease (with recurrent
respiratory infections), pancreatic insufficiency (which leads to malabsorption and growth
retardation) and elevated sweat electrolytes. tambm caracterizada por fezes com gordura
(esteatorreia) e obstruo intestinal.
The development of a disorder such as CF is greatly influenced by environmental factors and
genetic factors other than CFTR .
Nota: Defects in CFTR are also the cause of congenital bilateral absence of the vas deferens
(CBAVD). CBAVD is an important cause of sterility in men and could represent an incomplete
form of cystic fibrosis, as the majority of men suffering from cystic fibrosis lack the vas
deferens
O gene CFTR, transporte trans-membranar de Cl
-
e a clnica da fibrose qustica
O gene CFTR (cystic fibrosis transmembrane conductance regulator) est localizado em
7q31.2. A variao allica normal do gene de 250,000 pares de bases e contm 27 exes.
A protena codificada pelo gene CFTR corresponde a um transportador ABC (ATP-binding
cassette). CFTR transporta ies Cloro (Cl
-
) atravs da membrana celular dos pulmes,
fgado, pncreas, tracto digestivo e reproductivo e pele. CFTR endocitado da membrana
plasmtica por um processo dependente da clatrina que envolve dinamina e pela protena
AP2, que se liga clatrina. Vrios membros da famlia Rab de GTPases tambm controlam o
trfico de CFTR. The transport of chloride ions helps control the movement of water in tissues,
which is necessary for the production of thin, freely flowing mucus. Mucus is a slippery
substance that lubricates and protects the lining of the airways, digestive system, reproductive
system, and other organs and tissues.The structure of the complete CFTR protein has not yet
been experimentally determined. This is because membrane proteins, such as CFTR, with
substantial hydrophobic ("water-fearing") regions are extremely difficult to crystallize, and X-
ray crystallography can only be carried out on protein crystals. By comparing the CFTR
protein sequence with that of other known ABC transporters, models depicting the structure of
CFTR have been proposed. CFTR is made up of five domains: two membrane-spanning
domains (MSD1 and MSD2) that form the chloride ion channel, two nucleotide-binding
domains (NBD1 and NBD2) that bind and hydrolyze ATP (adenosine triphosphate), and a
regulatory (R) domain. CFTR is the only ABC transporter known to possess a regulatory
domain. Modification of the regulatory domain, either through the addition or removal of
chemical phosphate groups, has been shown to regulate the movement of chloride ions
Sebenta de Gentica
FCML
88
across the membrane. Os receptores |2- adrenrgico e 2b da adenosina, com adenilato-
ciclase (AC) e protena de ligao de GTP, activam CFTR.
Fig. 442: Diagram depicting the five domains of the CFTR membrane protein
A mutao F508 e as outras mutaes do gene CFTR. Heterozigotos compostos
Mutao F508: About 70% of mutations observed in CF patients result from deletion of three
base pairs in CFTR's nucleotide sequence- mutao missense. This deletion causes loss of
the amino acid phenylalanine located at position 508 in the protein; therefore, this mutation is
referred to as F508.
Fig. 45: The F508 deletion is the most common cause of CF. The isoleucine (Ile) at amino acid position 507
remains unchanged because both ATC and ATT code for isoleucine
F508 occurs in the DNA sequence that codes for the first nucleotide-binding domain (NBD1).
With normal CFTR, once the protein is synthesized, it is transported to the endoplasmic
reticulum (ER) and Golgi apparatus for additional processing before being integrated into the
cell membrane. When a CFTR protein with the F508 mutation reaches the ER, the quality-
control mechanism of this cellular component recognizes that the protein is folded incorrectly
and marks the defective protein for degradation. As a result, F508 never reaches the cell
membrane. People who are homozygous for F508 mutation tend to have the most severe
Sebenta de Gentica
FCML
89
symptoms of cystic fibrosis due to critical loss of chloride ion transport. This upsets the sodium
and chloride ion balance needed to maintain the normal, thin mucus layer that is easily
removed by cilia lining the lungs and other organs. The sodium and chloride ion imbalance
creates a thick, sticky mucus layer that cannot be removed by cilia and traps bacteria,
resulting in chronic infections. While the mechanism that leads to lung damage is not fully
understood, lung disease is the leading cause of morbidity and mortality among CF patients.
Tabela 1. Tipos de mutao mais comuns em CTFR
Outras mutaes: more than 900 mutations are known in the CFTR gene. As outras mutaes
mais encontradas no gene CFRT para alm da F508 foram: G542X (4,29%), R1162X
(2,14%), N1303K (1,43%) e R553X (0,71%) e G551D. Different CFTR mutations result in
different disease phenotypes. Some may have little or no effect on CFTR function, and some
may result in milder forms of disease. For example, one study in the Netherlands indicated
that CF patients who had one copy of F508 deletion and one of A455E mutation generally
expressed a milder form of pulmonary disease than those who were homozygous F508.
Also, a molecular physiologic analysis of the L441P mutation of CFTR revealed that the
L441P mutation completely abolished the CFTR Cl(-) channel activity by disrupting proper
protein folding and membrane trafficking of CFTR protein.
Fig. 46: Classes de mutaes na FC
Sebenta de Gentica
FCML
90
Nota: The presence of variant forms of genes other than CFTR can also affect disease
phenotype. Meconium ileus (MI), a severe intestinal obstruction, is observed in 15% to 20% of
babies born with cystic fibrosis. No CFTR gene mutations have been associated with MI. A
1999 study has shown that cystic fibrosis modifier 1 (CFM1), a modifier gene located on
chromosome 19, may determine MI susceptibility.
Heterozigotos compostos: gentipo em que esto presentes dois alelos mutantes diferentes,
no mesmo locus de cromossomas homlogos (ex. um indivduo com um alelo para a
hemoglobina S e um alelo para a hemoglobina C, o que origina hemoglobina SC).
A migrao e expanso europeia de alelos mutantes de CFTR
Cystic fibrosis (CF) is the most frequent severe recessive disorder in European populations.
The mutation spectrum of this disease, that is, the pool of alleles that cause CF in
homozygosis or in heterozygosis with another mutation, is made up of more than 1000
different mutations; of those, a three-basepair deletion, F508, accounts for roughly two-thirds
of cases, and only four others are found at average frequencies >1%.
The geographic distribution of mutation frequencies is heterogeneous among European
populations, showing a high geographic variation; F508 is ubiquitous, although its frequency
ranges from less than 50% to almost 100%; other mutations reach significant frequencies only
in part of the European continent, and many are rare and population specific. Most of the
knowledge about frequency distributions has been obtained on a mutation-by-mutation basis
and has consisted of mapping mutation frequencies, and the consequent association of their
geographic distribution with hypothetic demographic expansions of historical populations such
as the Celts or the Phoenicians. However, this single-mutation approach fails to capture the
complexity of both the mutation spectrum and the population history. Gene flow among
populations tends to disperse a number of mutations simultaneously and not just one.
Moreover, too often these ad hoc explanations are put forward without any quantitative
considerations about the extent of gene flow required or for the basic population genetics
involved. A different, sounder approach is possible: considering the whole mutation spectrum
at once and, beyond the mere description and subsequent story telling, applying to it the basic
toolkit of population genetics, which can lead to more accurate interpretations of the history of
the populations in relation to the natural history of genetic diseases. One basic requirement for
that approach is the reasonable assumption that all CF mutations are selectively equivalent.
That is, they cause the same loss of fitness in homozygosity and they give the same
advantage to heterozygotes. Under this assumption, Reich and Lander modelled mutation
diversity as a function of population history and of the overall frequency of disease-causing
Sebenta de Gentica
FCML
91
alleles. However, they considered one average spectrum per disease, and, allele diversity can
be very different among populations for the same disease. A basic property of a mutation set
is its frequency spectrum, that is, how many mutations are found and at frequency in a
particular population. It can be understood acumulatively, that is, how many mutations are
required to explain a given relative frequency (say 90%) of CF chromosomes. This has an
obvious interest in clinical testing, but it is also crucial in population genetics, since a
frequency spectrum may be the result of population history and of other evolutionary factors.
As for the direction of the clines, F508 peaks in NW Europe and declines towards SE
Europe, G542X declines from SW to NE Europe, G551D is almost restricted to NW Europe,
and N1303 K and W1282X show gradients from SW Asia and SE Europe towards NW Europe
(mais info: ver bibliografia Spatial patterns of cystic fibrosis mutation spectra in European
populations).
Fig. 47: Geographical distribution of F508 mutation in 94 middle Eastern, North African, and European
populations
Glocosuria renal Familiar. Mutao do gene SLC5A2. Mutaes privativas
Renal glucosuria is the excretion of glucose in the urine in detectable amounts at normal
blood glucose concentrations in the absence of any signs of general renal tubular dysfunction.
In general, renal glucosuria is a benign condition and does not require any specific therapy.
Glucosuria may be associated with tubular disorders such as Fanconi syndrome, cystinosis,
Wilson disease, hereditary tyrosinemia, or oculocerebrorenal osteodystrophy (Lowe
syndrome). Glucose is freely filtered by the glomerulus. Reabsorption of glucose
predominantly occurs on the brush border membrane of the convoluted segment of the
proximal tubule. Glucose enters the tubular cells by an active carrier-mediated transport
process, which is sodium-dependent, and exits via the basolateral membrane by facilitated
Sebenta de Gentica
FCML
92
diffusion by a glucose transporter, which is sodium-independent. The sodium-dependent
glucose cotransporters are a family of glucose transporters found in the intestinal mucosa of
the small intestine (SGLT1) and the proximal tubule of the nephron (SGLT2, predominantly,
and SGLT1). They contribute to renal glucose reabsorption. SGLT1 and SGLT2 are members
of the SLC5A gene family. SGLT2 is a low-affinity sodium/glucose cotransporter responsible
for the bulk of tubular reabsorption of filtered glucose. The SGLT2 sodium/glucose
cotransporter contains 672 amino acid residues and is almost exclusively expressed in the
kidney. SGLT1 contains 664 amino acid residues and is a high amino acid cotransporter
protein that is strongly expressed in the small intestine and, to some extent, in the kidney and
is responsible for reabsorption for the bulk of the remaining glucose. These 2 cotransporters
have significant homology; 59% of the amino acids are identical after alignment. Despite the
homology between the two, only one mutation is common: Arg137His. The human intestinal
SGLT1 has been localized to chromosome 22. SGLT1 is found in the straight segment of the
proximal tubule. The other cotransporter is SGLT2, which is expressed in the S1 segment of
the proximal tubule and is localized to chromosome 6. The facilitative glucose transporters
have isoforms GLUT 1-5. GLUT2 is mainly associated with glucose transport in the
convoluted portion of the proximal tubule. In segments with high reabsorptive rates (S1 and
S2 segments), the carrier is high capacity, low affinity. At birth, a high-affinity, low-capacity
pathway is also present to compensate for the reduced activity of the high-capacity, low-
affinity pathway. Familial renal glycosuria (FRG) is a rare renal tubular disorder caused by
mutations with the SLCA2 gene. This gene is mapped to chromosome 16p11.2. The mode of
inheritance that best fits FRG has been suggested to be codominance with incomplete
penetrance. Many heterozygous individuals display mild glucosuria (<10 g/1.73 m
2
/24 h),
whereas homozygous or compound heterozygous individuals usually have severe renal
glycosuria in excess of 10 g/1.73 m
2
/24 h.
Sndrome de Antley-Bixler. Mutao do gene POR
Cytochrome P450 oxidoreductase deficiency is a disorder of hormone production. This
condition specifically affects steroid hormones, which are needed for normal development and
reproduction. The hormonal changes associated with cytochrome P450 oxidoreductase
deficiency can affect the development of the reproductive system, skeleton, and other parts of
the body. This condition is inherited in an autosomal recessive pattern and the prevalence of
cytochrome P450 oxidoreductase deficiency is unknown. About 60 affected individuals have
been reported worldwide. The signs and symptoms of cytochrome P450 oxidoreductase
deficiency vary from mild to severe. Severe cases of cytochrome P450 oxidoreductase
deficiency are often given the name Antley-Bixler syndrome. The signs and symptoms of this
Sebenta de Gentica
FCML
93
condition are present at birth or become apparent in early childhood. Both males and females
with Antley-Bixler syndrome may have ambiguous genitalia or other genital abnormalities
caused by hormonal changes before birth. Hormonal changes may also disrupt sexual
development at puberty, leading to infertility. Antley-Bixler syndrome is also characterized by
skeletal abnormalities, particularly involving bones of the head and face. Other skeletal
abnormalities can include joint deformities (contractures) that limit movement, and
radiohumeral synostosis, which is a bone abnormality that locks the elbows in a bent position.
A blockage of the nasal passages (choanal atresia), intellectual disability, and delayed
development are also associated with Antley-Bixler syndrome. Cytochrome P450
oxidoreductase deficiency is caused by mutations in the POR gene (P450 (cytochrome)
oxidoreductase gene) on chromosome 7q11.2. This gene provides instructions for making the
enzyme cytochrome P450 oxidoreductase, which plays a critical role in the formation of
steroid hormones. This group of hormones includes testosterone and estrogen,
corticosteroids, (involved in the body's response to stress); and aldosterone, which helps
regulate the body's salt and water balance. Mutations in the POR gene reduce the activity of
cytochrome P450 oxidoreductase, which disrupts the production of steroid hormones.
Changes in sex hormones such as testosterone and estrogen lead to problems with sexual
development before birth and at puberty. It is less clear how impairment of cytochrome P450
oxidoreductase leads to skeletal abnormalities in people with Antley-Bixler syndrome. Studies
suggest that these abnormalities may be related, at least in part, to the abnormal production of
cholesterol. Cholesterol is a substance that is altered to produce steroid hormones. It is
necessary for normal embryonic development and is involved in bone formation. Although
Antley-Bixler syndrome is often caused by mutations in the POR gene, the characteristic
features of this disorder can also result from mutations in another gene, FGFR2. Some
researchers use the name Antley-Bixler syndrome to describe this pattern of features
regardless of the cause. However, others argue that the name should be reserved for severe
cases of cytochrome P450 oxidoreductase deficiency caused by POR gene mutations.
Hemoglobinopatias
Hemoglobina Lepore e crossing-over desigual
Certos pacientes com |-talassmia, moderada a intensa, apresentam uma cadeia no-o
incomum e que tem um segmento homlogo extremidade N-terminal de uma cadeia o
normal, seguida de um segmento homlogo extremidade C-terminal de uma cadeia |
normal, formando uma cadeia de fuso o|, denominada hemoglobina Lepore. Estas cadeias
de fuso so formadas por um processo de crossing-over desigual entre os genes o e |, mal
alinhados. Tal facto possvel, pois os dois genes so altamente homlogos e so contguos,
Sebenta de Gentica
FCML
94
no cromossoma 11. Formam-se dois genes hbridos: um novo gene contendo uma deleco,
correspondendo fuso de um fragmento do gene o com um fragmento do gene | gene
Lepore; um produto recproco contendo uma duplicao, com um gene o e um gene |
normais, mas tambm com um gene resultante da fuso de um segmento |, seguido de um
segmento o - gene anti-Lepore (no se expressa).
Talassmias-deleco, represso da transcrio ou traduo e anomalias de
splicing
As talassmias so um grupo heterogneo de doenas, caracterizadas pela incapacidade ou
reduo da sntese de uma cadeia o ou |. Em certos casos a falta de uma das cadeias pode
ser compensada pelo aumento de outras. Este facto reduz o tempo de vida dos eritrcitos,
devido deposio das cadeias na superfcie interna das membranas, tornando-as rgidas.
As hemcias tomam a forma de diancitos. Na o talassmia a sntese da cadeia o
(cromossoma 16) nula ou reduzida, afectando a formao das hemoglobinas fetal (Hb F) e
adulta (Hb A e Hb A2). o- Talassmia heterozigtica tipo I: 1 alelo mutado ou delectado; o
indivduo no apresenta sinais clnicos (portador silencioso) e produz 75% de cadeias o. o-
Talassmia heterozigtica tipo II: 2 alelos mutados ou delectados; o indivduo produz 50% de
cadeias o. Apresenta micropolicitmia e hipocromia. Hemoglobinose H: 3 alelos mutados ou
delectados; o indivduo apresenta anemia hemoltica crnica moderada. Produz 25% de
cadeias o, verificando-se um aumento da hemoglobina H (Hb H: ||||). o-Talassmia
homozigtica: no h produo de cadeias o (deleco ou mutao de todos os genes o); h
morte fetal por hidropsia (edema extenso por acumulao de lquidos em todo o corpo) e
encontram-se hemoglobinas de Barr (Hb de Barr: ). |-Talassmia: hemoglobinopatia que
resulta de uma diminuio na taxa de sntese das cadeias | (cromossoma 11) e afecta
apenas a sntese de hemoglobina A (Hb A). Deve-se normalmente a defeitos no gene | e no
a deleces. Tem um padro autossmico recessivo. Indivduos com dois alelos mutantes da
| globina apresentam Talassmia major (|-Talassmia homozigtica ou doena de Cooley).
Talassmia por deleco
De frequncia rara, resulta de uma deleco de um segmento varivel de DNA do |-cluster
(cromossoma 11), podendo envolver alm do gene | outros prximos (o, ). Se houver
deleco dos genes | e o, com manuteno do gene , temos como consequncia, a
persistncia hereditria da HbF - PHHF. Pode ocorrer tambm a deleco apenas do gene |,
com a manuteno do o, pelo que vai haver um alto nvel de HbA
2
. Se estas deleces forem
heterozigticas no h riscos clnicos nem patologia.
Sebenta de Gentica
FCML
95
Talassmia por represso da transcrio
Uma mutao na zona promotora impede o incio da transcrio.
Talassmia por represso da traduo
A mutao gera um codo de terminao que interrompe a sntese do polipptido.
Talassmia por anomalias do splicing
Uma mutao das sequncias de consenso, 5'GT, 3'AG, leva a alteraes dos locais de
"corte" e "emenda".
Talassmia por alterao da traduo
Uma mutao do 1 nucletido do mRNA (na transcrio) impede o acrscimo de 7-
metilguanosina, expondo assim o mRNA degradao enzimtica.
Drepanocitose e haplotipos de restrio
O defeito molecular consiste na existncia da hemoglobina falcmica (Hb S), que resulta da
substituio de cido glutmico por valina, na 6 posio da extremidade N-terminal da
cadeia |. O defeito primordial, corresponde a uma mutao pontual, na sequncia
nucleotdica do DNA (AT), no codo do gene |-globina HBB (cluster do cromossoma 11). O
produto da transcrio do codo vai ser GUG (valina), em vez do GAG original (glutamato).
uma doena autossmica recessiva, caracterizada por uma anemia hemoltica grave, em que
h tendncia das hemcias em assumir a forma de foice em condies de baixa tenso de
oxignio. Nos indivduos heterozigticos tanto o alelo mutado como o normal so
codominantes na expresso das protenas, por isso possuem HbS e HbA. A base fsica para
esta alterao da forma da hemoglobina a seguinte: a hemoglobina apresenta normalmente
glutamato na posio 6 da cadeia |, que se encontra em contacto com a gua e polar; a
sua substituio por uma valina, que apolar, cria uma ''zona aderente" na face externa da
molcula. Na superfcie da HbS desoxigenada existe um complemento da zona aderente que
fica oculto quando esta est oxigenada. Assim, a Hb forma agregados ou polmeros, que
precipitam dentro da hemcia, conferindo-lhe a sua forma caracterstica. As manifestaes
clnicas incluem anemia, ictercia e crises falcmicas associadas a obstruo vascular e
enfartes dolorosos em vrios tecidos (ossos, bao e pulmes). A drepanocitose resulta de um
estado de homozigotia; dos indivduos heterozigticos diz-se que possuem um trao
falcmico. Os indivduos heterozigticos no apresentam sintomas clnicos e possuem
hemoglobinas Hb A e Hb S. Estes indivduos apresentam uma vantagem adaptativa, sendo
mais resistentes infeco por Plasmodium falciporum (malria), da que exista uma
distribuio preferencial em zonas onde a malria endmica. A presena de um grande
Sebenta de Gentica
FCML
96
nmero de polimorfismos no locus | da hemoglobina tornou possvel o estudo da origem da
mutao, bem como a difuso das hemoglobinopatias correspondentes. Os resultados
parecem evidenciar que a mutao que condiciona a drepanocitose ocorreu pelo menos
quatro vezes em diferente reas geogrficas (Senegal, Benin, Repblica Aficana Central e
Arbia) e foi, depois difundida atravs da seleco pela existncia de malria nessas regies.
A HbS dos mediterrneos parece ser de origem africana e ambos os haplotipos originrios do
Benin e do Senegal foram tambm encontrados no mediterrneo.
Os polimorfismo de DNA no cluster do gene |
S
tem sido exaustivamente estudados usando a
tecnologia das endonucleases de restrio (antecedidas de PCR).
Diagnstico talassmias e drepanocitose:
The 1975 International Committee for Standardization in Hematology expert
panel on
abnormal Hbs and thalassemias made diagnostic recommendations
regarding the laboratory
investigation of these conditions.
Initial tests recommended include a complete blood count
(CBC),
electrophoresis at pH 9.2, tests for solubility and sickling, and
quantification of Hb A
2
and Hb F. If an abnormal Hb is identified
on the preliminary tests, then further techniques were
recommended
to identify the variants. These techniques include electrophoresis
at pH 6.06.2,
globin chain separation, and isoelectric
focusing (IEF- electrophoretic technique with excellent
resolution). Additional testing, including heat and isopropanol
stability tests, was
recommended for detection of unstable Hbs
or Hbs with altered oxygen affinity. After
presumptive identification of hemoglobinopathies and thalassemia
syndromes, and particularly
for purposes of genetic counseling,
defining the mutation or deletion present may be required.
Several
molecular techniques are available.
DNA from white blood cells, amniocytes, or
chorionic tissue
may be utilized for diagnosis of various - and -globin
chain abnormalities.
Typically, deletional mutations causing
-thalassemia syndromes and some rare -
thalassemias are
diagnosed using Southern blot hybridization of particular restriction
enzyme
digests to labeled complementary gene probes.
PCR techniques using allele-specific probes
after globin gene amplification,
allele-specific primers, or deletion-dependent amplification
with
flanking primers are used in definition of known globin
chain mutations/deletions, including
those for Hb S, E, D, and O,
and several -thalassemias. For unknown mutations, several
PCR-based methods, including
denaturing gradient gel electrophoresis, single-strand
conformation polymorphism
analysis, RFLP as well as sequencing of the amplified globin
gene
DNA may be used.
Origem e distribuio das mutaes nas hemoglobinopatias e gentica de populaes
Sebenta de Gentica
FCML
97
Hemoglobinopathies are especially common in areas in which malaria is endemic. This
clustering of hemoglobinopathies is assumed to reflect a selective survival advantage for the
abnormal red cells, which presumably provide a less hospitable environment during the
obligate intraerythrocytic stages of the parasitic life cycle. Very young children with
othalassemia are more susceptible to infection with the nonlethal Plasmodium vivax.
Thalassemia might then favor a natural protection against infection with the more lethal
P.falciparum. Thalassemias are the most common genetic disorders in the world, affecting
nearly 200 million people worldwide. About 15% of American blacks are silent carriers for
othalassemia; o-thalassemia trait (minor) occurs in 3% of American blacks and in 1to 15% of
persons of Mediterranean origin. | Thalassemia has a 10 to 15% incidence in individuals from
the Mediterranean and Southeast Asia and 0.8% in American blacks. The number of severe
cases of thalassemia in the United States is about 1000. Sickle cell disease is the most
common structural hemoglobinopathy occurring in heterozygous form in about 8% of
American blacks and in homozygous form in 1 in 400. Between 2 and 3% of American blacks
carry a hemoglobin C allele.
Fig. 49: Hb E e Hb S no Mundo
2. Patologia Gentica monognica autossmica dominante________
Anlise de mecanismos de expresso clnica autossmica dominante
Mecanismos em Dominantes:
Leses bsicas
Protenas estruturais
Sebenta de Gentica
FCML
98
Protenas de superfcie
Genes supressores de tumores
Fisiopatologia molecular
Perda de funo dominante (~ haploinsuficincia: ocorre quando uma reduo de 50% no
nvel da funo do gene causa um fentipo anromal)
Ganho de funo dominante (algumas mutaes criam um ganho de funo. Podem dar
origem: a um aumento de funo (por duplicao do gene, mutao activadora), a uma nova
funo (mutao especfica ou por formao de gene hbrido) ou a uma protena que se
deposite e se torne lesiva para a clula. Frequentemente, estas mutaes originam patologia
mesmo em heterozigotia, isto , do fentipos dominantes.
Efeito dominante negativo
Hipercolestrolmia familiar
A hipercolesterolemia familiar uma doena gentica associada a deficincia do
metabolismo lipdico, nomeadamente o transporte e metabolismo do colesterol. uma
doena com transmisso autossmica dominante com penetrncia de quase 100%. A
hipercolesterolemia familiar pode ser causada por mutaes em pelo menos trs genes
diferentes: receptor da LDL (LDLR) , APOB, e PCSK9. As mutaes no gene LDLR so
responsveis por cerca de 85% dos casos e as mutaes nos genes APOB e PCSK9
explicam em conjunto cerca de 5% dos casos. Em cerca de 10% dos casos ainda no foi
identificado o gene responsvel. O gene para o receptor LDL encontra-se no cromossoma
19p13.2 e contm 18 exes. As mutaes mais frequentes em cada um desses exes esto
na figura 50.
Fig. 50: Mutaes mais frequentes no gene do receptor LDL
Especifica os
21 a.a.do
pptido sinal
Sebenta de Gentica
FCML
99
NOTA: A famlia das protenas do receptor das LDL, tambm envolvidas na sinalizao Wnt.
Quanto ao gene que codifica a PCSK9 (protease proprotena convertase subtilisina/kexina
tipo 9), este reduz os nveis do receptor LDL no fgado de rato.
Variantes de PCSK9 associadas concentrao plasmtica de colesterol- LDL:
- Protena PCSK9 com os domnios de homologia ao prodomnio e ao domnio cataltico
da famlia das proprotenas convertase subtilisina.
- Variantes que co-segregam com hipercolesterolmia em famlias com hipercolesterolmia
familiar e variantes associadas a valores de colesterol-LDL abaixo do normal.
Fig. 53: Gene da PCSK9
Alteraes nos colagneos
Osteognese imperfeita e sindrome de Elhers-Danlos dominante
Osteognese imperfeita: mutaes nos genes COL1A1 ou COL1A2 (codificam o colagneo
tipo I) produzem osteognese imperfeita (doena dos ossos quebradios). A Osteognese
imperfeita (OI) tipo I a forma mais leve e causada por mutaes de mudana de fase e
mutaes sem sentido enquanto os tipo de OI mais graves so causados por substituies de
a.a. nas unidades repetidas Gly-X-Y
Sndrome de Elhers-Danlos: Resulta de defeitos na estrutura ou sntese deficiente do
colagneo por mutao no exo 6 do gene que o codifica (COLA1 e COLA2), ou em genes
de enzimas que participem na sua sntese. A natureza gentica desta doena heterognea,
havendo cerca de 10 variantes que podem ser dominantes ou recessivas: na herana
dominante, o gene estrutural do colagneo est alterado (se o indivduo for heterozigtico,
50% do colagneo normal); na herana recessiva, h uma deficiente maturao do
colagneo devido a uma alterao da enzima lisil transferase - herana ligada ao X. A
patologia expressa-se ao nvel do tecido conjuntivo, que fica instvel e laxo.
Sebenta de Gentica
FCML
100
A mutao da transtirretina e polineuropatia amiloidtica familiar (PAF)
Polineuropatia amiloidtica familiar (PAF): Uma amiloidose resulta da deposio de vrias
protenas fibrilhares insolveis nos tecidos, de tal modo que a sua funo afectada. As
fibrilhas representam, normalmente fragmentos proteolticos de vrias protenas plasmticas
e possuem uma estrutura em folha pregueada. O caso da polineuropatia amioidtica familiar
trata-se de um tipo de amiloidose congnita, autossmica dominante, resultante de uma
mutao do gene da transtirretina (TTR) localizado no cromossoma 18q11.2-q12.1 (4 exes,
o primeiro codifica para o pptido sinal). A TTR uma protena plasmtica (pr-albumina)
responsvel pelo transporte de retinol (retinal binding protein- RBP) e tiroxina (T4), sendo
sintetizada no fgado e crebro (metabolismo na fig. 54).
Fig. 54: Mecanismo da TTR
O gene ATTR apresenta 4 exes sendo que o primeiro exo codifica para o pptido sinal.
Uma mutao neste exo (Val20Ile) no atinge o pptido sinal. Mais de 70 das restantes
mutaes da TTR (heterogeneidade allica) ocorrem nos restantes 3 exes.
A variante portuguesa TTR (Met30) apresenta uma incidncia relativamente elevada na
Pvoa do Varzim (onde denominada doena dos pzinhos), sendo este o presumvel local
em que ter aparecido pela primeira vez esta variante- efeito fundador. H tambm focos
desta variante no Japo e na Sucia, porventura explicveis pela miscigenao portuguesa
nestes pases. A mutao TTR (Met30) condiciona uma menor solubilidade da protena
devido presena de um aminocido hidrofbico (metionina), pelo que esta se deposita
progressivamente na na banha dos nervos autnomos e perifricos, condicionando uma
alterao nas propriedades de conduo da membrana dos mesmos. Como a deposio
ocorre gradualmente e necessrio atingir uma acumulao significativa nos tecidos para
que a condio se manifeste, trata-se de uma condio com abiotrofia. Do mesmo modo se
pode explicar o facto de o gene que condiciona a doena ser dominante, visto que para que
ocorra a deposio da protena, que a causa primria da doena, basta que apenas um dos
Sebenta de Gentica
FCML
101
alelos condicione uma protena menos solvel. Os pacientes que sofrem polineuropatia
sensrio-motora (perifrica) apresentam uma diminuio da conduo neuro-muscular, com
incapacidade progressiva de estimulao nervosa, levando a atrofia da musculatura lisa e
estriada e, logo, diminuio acentuada da capacidade de locomoo e motilidade gastro-
intestinal.
O sndrome de Marfan e a fibrilhina
A sndroma de Marfan uma doena relativamente frequente, transmitida como autossmica
dominante e que afecta o tecido conjuntivo. Os seus sinais clnicos afectam o olho e os
sistemas esqueltico e cardiovascular e incluem a deslocao do cristalino (ectopia lentis),
aracnodactilia, hiperextensibilidade das articulaes e fragilidade da mdia da aorta, levando
dilatao da croa da aorta. Pensa-se que Araham Lincoln poder ter sofrido desta doena.
A maioria dos casos devida a uma mutao (normalmente do tipo missense) no gene para
a fibrilina, localizado no brao longo do cromossoma 15. A fibrilina uma glicoprotena de
elevada massa molecular (350 kDa), sendo o constituinte estrutural das microfibrilhas de 10 -
12 nm presentes na maior parte dos tecidos conjuntivos. produzida nos fibroblastos, sendo,
aps clivagem proteoltica, segregada para o meio extracelular onde incorporada a
microfibrilhas insolveis, as quais se pensa que formam uma matriz de suporte para a
deposio de fibras de elastina. De especial relevncia para esta condio a distribuio da
fibrilina nos tecidos, sendo encontrada em maior abundncia no ligamento suspensor do
cristalino, no peristio e associada s fibras de elastina na aorta, o que explica os sinais
clnicos caractersticos j apontados. Outras protenas esto tambm presentes nas
microfibrilhas e parece provvel que anomalias estruturais que as afectem possam causar
esta mesma condio (ou outras), pelo que a sndroma de Marfan um caso em que poder
existir (real ou potencialmente) heterogeneidade gentica.
Correia de Huntington
Doena do sistema nervoso central (SNC), autossmica dominante, com penetrncia
completa que se caracterizada pela perda da capacidade motora e intelectual, associada a
tremores e demncia. Foi o primeiro distrbio mendeliano a ser mapeado, por ligao a
marcadores de DNA. O defeito gentico bsico permanece desconhecido, mas sabe-se que
est localizado no brao curto do cromossoma 4 (4p16). Se o gene for herdado da me,
manifesta-se por volta dos 70 anos (por vezes 50 ou 60); se for herdado do pai, manifesta-se
mais cedo (40 anos) e com expresso mais grave, o que leva a supor a existncia de um
fenmeno de imprinting. Caso tpico de abiotrofia. No tem tratamento especfico.
Sebenta de Gentica
FCML
102
Caractersticas do gene responsvel pela doena de Huntington
- Codifica uma protena (huntingtina), de aproximadamente 348kDa, cuja funo ainda no
foi totalmente identificada (est associada aos microtbulos nas dendrites e extremidades
dos axnios)
- Apresenta repeties do tripleto CAG no exo 1, codificando glutamina, na regio 5 do
gene normal (wild type), dentro da regio codificadora da protena presuposta;
- As repeties de CAG formam poliglutamina que constitui estruturas | no removidas por
chaperones.
- Os genes de indivduos normais apresentam 6 a 35repeties do tripleto CAG; os de
individuos afectados pela doena apresentam mais do que 35 repeties; indivduos com
HD de manifestao precoce apresentam + de 60 repeties do tripleto.
- Esta mutao um exemplo de uma expanso por repetio de um trinucletido;
- Aumento do comprimento da expanso do trinucletido est relacionado com uma
manifestao precoce da doena de gerao para gerao: antecipao.
Neurofibromatose de von Recklinghausen
Resulta de um defeito gentico bsico no cromossoma 17 que consiste numa mutao do
gene NF1 (com um intro partilhado com 3 outros genes, os quais podero afectar as
manifestaes clnicas dos doentes com NF1), presumivelmente, um gene supressor de
tumor com perda somtica no desenvolvimento de alguns tumores, visto que foi demonstrado
que nesta doena os tumores tm uma origem policlonal, sugerindo, deste modo, que cada
clula possui uma tendncia proliferativa intrnseca. Este gene NF1 apresenta alta taxa de
neo-mutaes, sendo de novo cerca de 50% das mutaes. Nos restantes casos, a
severidade do quadro clnico parece estar associada transmisso por via materna. H
casos descritos em que os indivduos afectados apresentavam mosaicismo somtico. A
Protena codificada por NF1, a neurofibromina, , possvelmente, activadora de GTPase
regulando a funo do Ras (inactivando-o) e participando no controlo da diviso celular. A
heterogeneidade da regulao pos-transcripcional da expresso de NF1 parece tambm ser
importante uma vez que um estudo demonstrou que alguns pacientes com Neurofibromatose
apresentam cerca de 3 a 12% de edio C-U do RNA.
Sebenta de Gentica
FCML
103
Fig. 55: Protena NF activadno a Ras
Existe apenas tratamento clnico ou cirrgico para os defeitos secundrios. Afecta cerca de 1
em 3000 indivduos. Caso tpico de expressividade varivel, com penetrncia completa (90 a
100%). As manifestaes da doena podem variar de um espectro completo:
- Muitos tumores benignos subcutneos (fibromatomas), sendo frequente a sua
transformao maligna;
- Manchas "caf au lait" ;
- Atraso mental;
- Hematomas pigmentados na ris (ndulos de Lish);
- Manifestaes sseas (escoliose) e intracranianas (meningoblastoma, etc);
- Manifestaes endcrinas (puberdade precoce, atrasos no crescimento)
Ou um espectro parcial, com apenas algumas manchas, que podem passar despercebidas.
Tais variaes, podem ocorrer, mesmo dentro da mesma famlia. A variabilidade de
manifestaes clnicas pode ser de atribuir ao splicing alternativo complexo e ao editing C-
U no mRNA.
A cardiomiopatia hipertrfica familiar e heterogeneidade de locus e allica
Cardiomiopatia hipertrfica (CMH): doena do miocrdio caracterizada por hipertrofia
ventricular esquerda, com um envolvimento predominante do septo interventricular. Tem uma
grande variabilidade inter e intra-familiar, variando entre formas benignas e formas malignas
com um elevado risco de insuficincia cardaca e morte sbita. uma doena de
hereditariedade autossmica dominante, com uma incidncia de 1:500-1.000. Em cerca de
55% dos casos h histria familiar evidente. Esta
patologia est ligada, entre outras anormalidades, a uma mutao gentica na cadeia de
betamiosina pesada- cMHC (gene MYH7 no cromossoma 14q1, freqncia de 35 a 50%).
Sebenta de Gentica
FCML
104
Vrios pesquisadores tm demonstrado que um nmero diferente de mutaes nas protenas
contrteis responsvel pela CH e age reduzindo a performance da contratilidade
miocrdica. Certos autores descrevem outras as alteraes de genes: miosina cardaca
(gene MYBPC no cromossoma 11q1, freqncia de 15 a 20%); troponina T cardaca (gene
TNNT2 no cromossoma 1q3, freqncia de 15 a 20%); -tropomiosina (gene TPM1 no
cromossoma 15q2, freqncia menor que 5%); troponina cardaca I (gene TNN3 no
cromossoma 19q13, freqncia menor que 1%). Alm dessas, j foram descritos: MLC1,
MLC2, actina -cardaca e um gene desconhecido localizado no cromossoma 7q3.
Diversos pathways em cascata, como alteraes energticas mitocondriais, tono simptico,
enzima conversora da angiotensina, frmacos, polimorfismos, podem conduzir/agravar o
fenotipo.
Heterogeneidade de locus: Condio em que um fentipo (ou fentipos muito semelhantes)
so determinados, de um modo independente, por alelos localizados em loci diferentes.
Heterogeneidade allica: Condio em que um fentipo ou fentipos muito semelhantes so
determinados, de um modo independente, por alelos diferentes localizados no mesmo locus.
Frequentemente, uma patologia pode ser provocada por diferentes tipos de mutaes num
gene heterogeneidade allica
Fig. 56: Heterogeneidade de lcus e allica
Heterogeneidade em CMH:
gentica allica clnica prognstico
Sebenta de Gentica
FCML
105
Doena poliqustica renal
Insuficincia renal crnica terminal com necessidade de dilise/transplante em 50% dos
doentes (57-73 anos). Polycystic kidney disease is a disorder that affects the kidneys and
other organs. The two major forms of polycystic kidney disease are distinguished by the usual
age of onset and their pattern of inheritance. The autosomal dominant form (sometimes called
ADPKD) has signs and symptoms that typically begin in adulthood, although cysts in the
kidney are often present from childhood. Autosomal dominant polycystic kidney disease can
be further divided into type 1 and type 2, depending on which gene is mutated. The autosomal
recessive form of polycystic kidney disease (sometimes called ARPKD) is much rarer and is
often lethal early in life. The signs and symptoms of this condition are usually apparent at birth
or in early infancy. Mutations in the PKD1, PKD2, and PKHD1 genes cause polycystic kidney
disease. Mutations in either the PKD1 or PKD2 gene can cause autosomal dominant
polycystic kidney disease. These genes provide instructions for making proteins whose
functions are not fully understood. Researchers believe that they are involved in transmitting
chemical signals from outside the cell to the cell's nucleus. The two proteins work together to
promote normal kidney development, organization, and function. Mutations in the PKD1 or
PKD2 gene lead to the formation of thousands of cysts, which disrupt the normal functions of
the kidneys and other organs. People with mutations in the PKD2 gene, particularly women,
typically have a less severe form of the disease than people with PKD1 mutations. The signs
and symptoms, including a decline in kidney function, tend to appear later in adulthood in
people with a PKD2 mutation. Mutations in the PKHD1 gene cause autosomal recessive
polycystic kidney disease. This gene provides instructions for making a protein whose exact
function is unknown; however, the protein likely transmits chemical signals from outside the
cell to the cell nucleus. Researchers have not determined how mutations in the PKHD1 gene
lead to the formation of numerous cysts characteristic of polycystic kidney disease. Although
polycystic kidney disease is usually a genetic disorder, a small percentage of cases are not
caused by gene mutations. These nonhereditary cases are called acquired polycystic kidney
disease. This form of the disorder occurs most often in people who have been treated for
several years with hemodialysis (a procedure that filters the blood in people with kidney
failure).
Clonagem dos genes responsveis por ADPKD
clonagem posicional PKD1:
- PKD1 ligado ao locus a-globina; anlise de linkage detalhada; carta de restrio do locus
PKD1; intervalo crtico em 16p13.3 posicionado entre GGG e SM7: 600 Kb, CpG rich;
identificao do transcripto
mecanismos genticos na ADPKD
Sebenta de Gentica
FCML
106
- estudos de perda de heterozigotia (LOH): a inactivao somtica do alelo normal
transforma o estatuto dominante em recessivo ao nvel celular.
outros genes responsveis por DQR
NPHP 1-6, Glis2, AH1 e PKHD1.
a hiptese ciliar (o clio tem funes no desenvolvimento do nefrnio e o modelo two-hit
explica a disfuno ciliar)
- As cistoproteinas (CP) so expressas primariamente no clio, corpo basal ou centrossoma
- As CP esto filogeneticamente conservadas e interagem maioritariamente entre si
- Os diferentes fentipos clnicos reflectem especificidade na expresso tissular de cada CP
- Funes sensoriais normais: fluxo urinrio tubular Ca
++
epitlio no proliferativo e
diferenciado
- Regulao do ciclo celular:
interfase: centrossoma centro organizao microtubular da clula
mitose: desaparecimento do clio e centrossoma centrolos fuso mittico
Retinopatia Pigmentar:
Quadro: Padres de Hereditariedade da RP
Pode ainda ser mitocondrial e dignica (forma infrequente em que a alterao gnica aparece
em dois genes diferentes num mesmo indivduo e s se manifesta caso os dois genes
apresentem mutao.)
Foram mapeados vrios genes envolvidos nesta patologia (mais de 100)
3. Genopatias Metablicas_____________________________________
Doenas lisossomais - mucopolissacaridoses e esfingolipidoses
As doenas lisossomais caracterizam-se por um defeito gentico nas enzimas hidrolticas dos
lisossomas, ocorrendo uma acumulao de determinadas substncias, que em condies
normais seriam degradadas pelas enzimas alteradas. Podem ser devidas falta de sntese
Sebenta de Gentica
FCML
107
das enzimas, ou falha dos mecanismos que introduzem as enzimas nos lisossomas. A
acumulao de substratos nos lisossomas leva disfuno e morte celular, assim como ao
aumento da massa dos tecidos e rgos onde esses produtos seriam degradados. Uma
caracterstica proeminente destas patologias a sua heterogeneidade gentica e clnica.
Mucopolissacaridoses
As mucopolissacaridoses so um grupo heterogneo de doenas de armazenamento, que
resultam da acumulao de mucopolissacridos parcialmente degradados, devido a um
defeito enzimtico geneticamente determinado em enzimas lisossomais especficas
envolvidas na degradao dos mucopolissacridos. Os mucopolissacridos acumulados so:
dermatano sulfato, heparano sulfato, keratano sulfato e condroitina sulfato. As enzimas
envolvidas em cada MPS clivam resduos glicdicos terminais das cadeias polissacardicas
dispostas ao longo de um polipeptdeo ou core proteico. Quando existe um bloqueio na
remoo de um acar terminal, a restante cadeia polissacrida no mais degradada e
estas cadeias acumulam-se nos lisossomas, em vrios tecidos e orgos. Resultam em
alteraes somticas e neurolgicas graves.
Sndrome
ManifestaesClnicas
DficeEnzimtico.
Mucopolissacrido
Acumulado/Excretado
Gentica
Hurler
Turvao da crnea, alteraes
esquelticas, aspectos faciais
grosseiros, articulaes rigdas,
atrasomental
oLiduronidase.
Sulfato de dermatano e
Sulfatodeheparano
Autossmica
recessiva;
cr.4
Scheie
Incio aps os 5 anos de idade.
Turvao da crnea,
comprometimento da viso, ,
inteligncia e expectativa de vida
normal,cardiopatiavascular
oLiduronidase.
Sulfato de dermatano e
Sulfatodeheparano
Autossmica
recessiva;
cr.4
Hurler/
Scheie
Fentipo intermdio entre as duas
sndromes.Tematrasomental.
oLiduronidase.
Sulfato de dermatano e
Sulfatodeheparano
Autossmica
recessiva;
cr.4
Hunter
Semelhantes s da sndroma de
Hurler mas com progresso mais
lenta, ausncia da turvao da
crnea, leso cutnea "seixosa"
peculiar
Iduronatosulfatase.
Sulfato de dermatano e
Sulfatodeheparano
Autossmica
recessiva ligada
aoX
SanfilippoA
Hiperactividade, atraso mental
severo, neurodegenerao
progressiva.
HeparanNsulfatase. Sulfato
deheparano
Autossmica
recessiva
SanfillipoB semelhantes s da sndrome do tipo
A
oNacetilglucosaminidase.
Sulfatodeheparano
Autossmica
recessiva
Quadro 7: Mucolipossacaridoses
Sebenta de Gentica
FCML
108
Esfingolipidoses
Os esfingolpidos so normalmente degradados nos lisossomas das clulas fagocticas,
(essencialmente macrfagos) do sistema reticuloendotelial, localizado no fgado, bao e
medula ssea especialmente no perodo neonatal. A via catablica dos esfingolpidos envolve
a aco de uma srie de enzimas que clivam ligaes especficas nos compostos: o e |-
galactosidases, |-glucosidase, neuraminidase, hexosaminidase, esfingomielinase
(fosfodiesterase esfingomielina-especfica), sulfato esterase (sulfatase), ceramidase (amidase
ceramida-especfica). Todas estas enzimas so glicoprotenas e esto contidas nos
lisossomas firmemente ligados sua membrana, sendo o seu pH ptimo de 3,5 a 5,5 - os
enzimas so hidrolases e, consequentemente, um dos substratos destas reaces a gua.
A maioria destas enzimas so relativamente estveis e surgem como isoenzimas. Quando a
actividade de uma das enzimas desta via marcadamente reduzida nos tecidos de um
indivduo, devido a uma mutao, o substrato da enzima ausente ou defeituosa acumula-se e
depositado nos lisossomas do tecido responsvel pelo catabolismo deste lpido; a
manifestao patolgica deste fenmeno corresponde s doenas de armazenamento
lipdico, das quais fazem parte as esfingolipidoses. As doenas de armazenamento de
esfingolpidos so, geralmente, recessivas e ocorrem apenas em homozigticos com uma
deficincia em ambos os cromossomas, Em todas estas doenas o alelo defeituoso encontra-
se num cromossoma autossmico, excepto na Doena de Fabry onde est ligado ao
cromossoma X.
Caractersticas comuns das esfingolipidoses:
- Geralmente apenas 1 esfingolpido se acumula nos rgos envolvidos
- A poro ceramida comum aos vrios lpidos armazenado
- A taxa de biossntese do lpido acumulado normal
- Uma enzima catablica encontra-se ausente em cada uma destas doenas
- A dimenso da deficincia enzimtica igual em todos os tecidos afectados
Quadro 8: Esfingolipidoses
Sebenta de Gentica
FCML
109
A doena de Tay-Sachs e a endogamia
Doena lisossmica de armazenamento, do tipo das gangliosidoses (esfingolipidose), na qual
a substncia acumulada o ganglisido (GM
2
). autossmica recessiva e caracteriza-se
pela carncia de Hexosaminidase A, responsvel pela clivagem do GM
2
. A HexA
cataliticamente activa o produto de trs genes que codificam as subunidades o e | da
enzima e uma protena activadora. Na doena de Tay-Sachs ocorre uma alterao no gene
que codifica a subunidade o do cromossoma 15). Na ausncia de HexA vai ocorrer uma
acumulao de GM
2
, principalmente ao nvel do crebro, levando ao fentipo de atraso
mental, cegueira progressiva e degenerescncia psicomotora. Se a mutao afectar o locus
da subunidade | do cromossoma 5) surge a Doena de Sandhoff, em que ocorre uma
deficincia em hexosaminidase A e B.
Like many genetic disorders, TSD is concentrated in specific communities defined by
endogamous cultural and ethnic patterns. Historically, Eastern European people of Jewish
descent (Ashkenazi Jews) have a high incidence of Tay-Sachs. Documentation of Tay-Sachs
in this Jewish population reaches back to 15th century Europe. In the United States, about 1
in 27 to 1 in 30 Ashkenazi Jews is a recessive carrier. Some theories have been proposed to
explain the high frequency of Tay-Sachs carriers in the Ashkenazi Jewish population:
- Heterozygote advantage. When applied to a particular allele, this theory posits that
carriers of the mutation have a selective advantage, perhaps in a particular
environment.
- Reproductive compensation. Parents who lose a child because of disease tend to
"compensate" by having additional children to replace them. This may maintain and
possibly even increase the incidence of autosomal recessive disease.
- Founder effect. This hypothesis states that the high incidence of the 1278insTATC
mutation is the result of random genetic drift, which amplified a high frequency that
existed by chance in an early founder population.
Sndromes de Hurler, Hunter e I cell disease - anlise dos mecanismos
etiopatognicos
Sndrome de Hurler
Mucopolysaccharidosis type I (MPS I) is a condition that affects many parts of the body. This
disorder was once divided into three separate syndromes, Hurler syndrome (MPS I-H), Hurler-
Scheie syndrome (MPS I-H/S), and Scheie syndrome (MPS I-S), listed from most to least
severe. Because there is no clear distinction between these three syndromes, MPS I is
Sebenta de Gentica
FCML
110
currently divided into the severe and attenuated types. Mutations in the IDUA (iduronidase,
alpha-L) gene cause MPS I. The IDUA gene provides instructions for producing an enzyme
that is involved in the breakdown of large sugar molecules called glycosaminoglycans
(GAGs). GAGs were originally called mucopolysaccharides, which is where this condition gets
its name. Mutations in the IDUA gene reduce or completely eliminate the function of the IDUA
enzyme. The lack of IDUA enzyme activity leads to the accumulation of GAGs within cells,
specifically inside the lysosomes. The accumulation of GAGs increases the size of the
lysosomes, which is why many tissues and organs are enlarged in this disorder. Researchers
believe that the GAGs may also interfere with the functions of other proteins inside the
lysosomes and disrupt the movement of molecules inside the cell.
Sndrome de Hunter
Mutations in the IDS (iduronate 2-sulfatase) gene cause MPS II (Mucopolysaccharidosis type
II) also known as Hunters syndrome. The IDS gene provides instructions for producing the
I2S enzyme, which is involved in the breakdown of large sugar molecules called
glycosaminoglycans (GAGs). GAGs were originally called mucopolysaccharides, which is
where this condition gets its name. Mutations in the IDS gene reduce or completely eliminate
the function of the I2S enzyme. Lack of I2S enzyme activity leads to the accumulation of
GAGs within cells, specifically inside the lysosomes. Lysosomes are compartments in the cell
that digest and recycle different types of molecules. Conditions that cause molecules to build
up inside the lysosomes, including MPS II, are called lysosomal storage disorders. The
accumulation of GAGs increases the size of the lysosomes, which is why many tissues and
organs are enlarged in this disorder. Researchers believe that the GAGs may also interfere
with the functions of other proteins inside the lysosomes and disrupt the movement of
molecules inside the cell.
Mucolipidose II (I-cell disease)
A mucolipidose II, ou I-cell disease (inclusion cell disease) uma doena rara autossmica
recessiva que resulta de uma topognese
*
disfuncional das enzimas lisossomais nas clulas
de origem mesenquimatosa. Esta condio deve-se deficincia da enzima N-acetil-
glucosamina fosfotransferase (GlcNAc), cujo locus foi mapeado em 4q21-q23. No entanto,
outros estudos genticos indicaram que mais do um produto gnico contribui para a
constituio da enzima funcional, mas nem cDNA nem o gene para a enzima foram clonados
at hoje. A sua deficincia condiciona um bloqueio na via que leva formao do sinal de
manose-6-fosfato (M6P), que reconhecido pelos receptores responsveis pela segregao
das enzimas lisossomais na rede trans-Golgi para os pr-lisossomas. Como tal, as enzimas
Sebenta de Gentica
FCML
111
lisossomais so canalizadas para a via de secreo e libertadas no meio extracelular.
Geralmente, os sinais da doena apresentam-se nascena e progridem gradualmente,
sobrevindo a morte durante a primeira dcada de vida, normalmente devido a falncia
cardaca ou na sequncia de graves infeces respiratrias. No existe ainda um tratamento
especfico e efectivo, sendo este essencialmente sintomtico.
Terapia gnica nas doenas de Fabry, de Gaucher e no sndrome de Hurler
Quadro 8: TRE: terapia de reposio enzimtica, uma modalidade de tratamento que consiste na aplicao
peridica duma enzima artificial, semelhante que o paciente no produz em quantidade suficiente e cuja
deficincia a responsvel pelo quadro clnico da doena. ISS: inibio da sntese do substrato
Sndrome de Fabry:
Alternative treatments for many lysosomal storage diseases have been developed, including
allogeneic organ and bone marrow transplantation, enzyme replacement therapy, and gene
therapy. Significant clinical risks still exist with allogeneic transplantations. -Gal A enzyme
replacement therapy has been implemented in clinical trials. This approach has been effective
but may have limitations for long-term systemic or cost-effective correction. As an alternative,
gene therapy approaches, involving a variety of gene delivery systems, have been pursued for
the amelioration of Fabry disease. Fabry disease is a compelling disorder for gene therapy, as
target cells are readily accessible and relatively low levels of enzyme correction may suffice to
reduce storage. Importantly, metabolic cooperativity effects are also manifested in Fabry
disease, wherein corrected cells secrete -Gal A that can correct bystander cells. In addition,
a broad therapeutic window probably exists, and mouse models of Fabry disease have been
generated to assist studies. As an example, in vitro and in vivo studies using -Gal A-
transduced haematopoietic cells from Fabry mice have demonstrated enzymatic correction of
recipient cells and dissemination of-Gal A upon transplantation, leading to reduced lipid
storage in a number of clinically relevant organs. This corrective enzymatic effect has recently
Sebenta de Gentica
FCML
112
been shown to be even further enhanced upon pre-selection of therapeutically transduced
cells prior to transplantation.
Sndrome de Gaucher:
The first of the two Gaucher disease treatments based on this strategy is allogeneic bone
marrow transplantation, which results in the repopulation of affected tissues with enzyme-
competent macrophages. The second approach to treatment which has resulted in clinical
improvement in Gaucher disease patients is macrophage-targeted enzyme replacement. This
treatment takes advantage of naturally occurring mannose receptors on macrophages and the
exposition of accessible mannose receptors in the oligosaccharides of glucocerebrosidase to
efficiently deliver the enzyme to macrophages. While both of these approaches to treating
Gaucher disease are important because they provide some means of therapy where none
previously existed, both approaches have significant limitations. Allogeneic bone marrow
transplantation has associated with it morbidity and mortality risks that are unacceptable for
many patients. Further, HLA matched bone marrow donors do not exist for the majority of
patients. As for macrophage-targeted enzyme replacement, it is currently an expensive and
life-long therapy; thus, it should be reserved for only the most severely ill patients. Despite the
limitations of these two therapies, their successes have demonstrated that enzymatic
correction of only one cell type, the macrophage, results in effective therapy for Gaucher
disease. From the point of view of developing somatic cell gene therapy for Gaucher disease,
the fact that marrow transplantation is effective demonstrates that bone marrow stem cells are
an appropriate target cell to which to transfer a "therapeutic" gene. Further, because of the
pivotal role of macrophages in Gaucher disease, alternative target cells to be considered for
gene transfer are the committed macrophage precursors, peripheral blood monocytes, or
cultures of bone marrow which are capable of producing macrophage precursors.
Sndrome de Hurler:
Allogeneic bone marrow transplantation is the most effective treatment for Hurler's syndrome.
However, due to a lack of matched related donors and unacceptable morbidity of matched
unrelated transplants, this therapy is not available to all patients. Therefore we have been
developing an alternative approach based on transfer and expression of the normal gene in
autologous bone marrow. A retroviral vector carrying the full length cDNA for alpha-L-
iduronidase has been constructed and used to transduce bone marrow from patients with this
disorder. A number of different gene transfer protocols have been assessed including the
effect of intensive schedules of exposure of bone marrow to viral supernatant and the
influence of growth factors. With these protocols we have demonstrated successful gene
Sebenta de Gentica
FCML
113
transfer into primitive CD34+ cells and subsequent enzyme expression in their maturing
progeny. Also, using long-term bone marrow cultures, we have demonstrated high levels of
enzyme expression sustained for several months. The efficiency of gene transfer has been
assessed by PCR analysis of haemopoietic colonies as around 50%. No advantage has been
demonstrated for the addition of growth factors or intensive viral exposure schedules. Indeed
a possible disadvantage has been identified for the use of intensive transduction procedures.
The enzyme is secreted into the medium and functional localisation has been demonstrated
by reversal of the phenotypic effects of lysosomal storage in macrophages. This pre-clinical
work forms the basis for a clinical trial of gene therapy for Hurler syndrome.
As vias metablicas da fenilalanina e tirosina - fenilcetonria, alcaptonria e
albinismo
Fenilcetonria (PKU): Doena de transmisso hereditria autossmica recessiva,
caracterizada por uma deficincia no metabolismo da fenilalanina. A sua frequncia de 1
para 13500-15000 nascimentos. Normalmente a fenilalanina hidroxilada a tirosina por
intermdio de uma reaco catalisada pela enzima fenilalanina hidroxilase, que tem como
cofactor a BH
4
(tetrahidrobiopterina). A fenilcetonria resulta de uma deficincia da enzima
(98% dos casos), ou do seu cofactor (2% dos casos). O gene PAH que codifica a enzima
encontra-se no brao longo do cromossoma 12 (12q). Com a acumulao da fenilalanina nos
lquidos corporais surgem vias alternativas, havendo sntese de c. fenilpirvico, c.
fenilactico e fenillctico. As manifestaes clnicas devem-se, por um lado, ao efeito txico
destes ltimos compostos e por outro lado, falta de neurotransmissores (dopamina,
noradrenalina e adrenalina) e melanina, derivados da tirosina. Estas manifestaes
consistem em deteriorao neurolgica ps-natal (oligofrenia fenilpirvica - QI <50), atraso
mental profundo, atraso no crescimento, agitao, agressividade, convulses, fraca
pigmentao da pele e cabelo claro e rashs cutneos eczematosos.
PKU apresenta uma causa multifactorial: por um lado implica uma mutao no gene PAH
(factor gentico) mas tambm uma exposio a uma dieta de fenilalanina (factor ambiental)
Alcaptonria: doena autossmica recessiva rara, provocada pela deficincia da enzima
cido homogentsico-oxidase, responsvel pela degradao do cido homogentsico
(alcaptona), no rim e fgado. Este um produto intermedirio do metabolismo da tirosina,
que, em consequncia da sua degradao incompleta, surge na urina, tornando-a escura. O
pigmento tambm se deposita noutros tecidos, como a cartilagem das orelhas e as
articulaes (ocronose), podendo nestas ltimas levar ao aparecimento de artrites.
Sebenta de Gentica
FCML
114
Albinismo: Doena autossmica recessiva causada pela falta de tirosinase, enzima que
catalisa a formao de melanina a partir da tirosina (oxidao da tirosina). O albinismo
apresenta heterogeneidade gentica, isto , as causas que conduzem falta de melanina
podem ter diversas origens, sendo provocadas por alteraes de uma ou mais enzimas, cujos
genes esto em diferentes cromossomas. O quadro clnico inclui cabelos brancos, pele
esbranquiada, ris rosada, hipersensibilidade luz solar. O albinismo ocular manifesta-se
como uma doena autossmica recessiva, ou tambm ligada ao X, embora o defeito
metablico permanea desconhecido a apresentao clnica consiste em baixos nveis de
melanina, escassa pigmentao do olho e intolerncia luz.
Deficincia de alfa-1-antitripsina
A o-1-antiproteinase (alfa-1-antitripsina) uma glicoprotena srica, constituda por uma
cadeia polipeptdica e trs cadeias de oligossacridos. Constitui mais de 90% da fraco o
1
das protenas do soro e sintetizada pelo fgado e macrfagos. Trata-se do principal inibidor
de vrias enzimas proteolticas tais como a tripsina, quimiotripsina e elastase, pois tem a
capacidade de formar complexos com estas enzimas. O seu locus, localizado no
cromossoma 14, altamente polimrfico com cerca de 50 alelos relativamente comuns e
cerca de 25 alelos raros. Cada alelo d origem a uma variante da enzima, com uma
actividade enzimtica especfica, sendo, o alelo M o mais vulgar (o gentipo MM condiciona o
produto fenotpico PiM). Em relao a esta protena h duas reas de interesse clnico. Uma
deficincia desta protena responsvel por 5% das doenas pulmonares obstrutivas
crnicas (DPOC). Um tipo muito especfico de enfisema familiar que se instala no jovem
adulto ocorre em indivduos que sintetizam a protenas PiZ (condicionada pelo gentipo ZZ),
a qual segregada em muito menor quantidade em relao PiM (na prtica, embora a
expresso dos alelos apresente codominncia, a deficincia em o1-antiproteinase uma
doena autossmica recessiva). Neste caso, a quantidade de o1-antiproteinase insuficiente
e quando ocorre a acumulao de neutrfilos no pulmo (ex.: durante uma pneumonia), o
indivduo afectado no possui um mecanismo eficaz para impedir a leso do parnquima
pulmonar pelas proteases, como a elastase, que so produzidas por estas clulas. De
particular interesse que um resduo de metionina desta protenas (resduo 358), que est
intimamente envolvido na formao do complexo com as proteases pode ser oxidado pelo
fumo do tabaco transformando-se em sulfxido de metionina, o que inactiva totalmente a
protenas. Este efeito particularmente devastador em indivduos que possuem o gentipo
ZZ, visto que j tinham os nveis de o1-antiproteinase muito baixos. Deste modo, a instalao
do enfisema pulmonar pode ser acelerada por um factor comportamental, como a exposio
ao fumo do tabaco (fenmeno marcadamente ecogentico). Neste caso, a determinao do
Sebenta de Gentica
FCML
115
gentipo de um paciente com enfisema pulmonar de extrema relevncia, visto que a
administrao endovenosa de o1-antiproteinase tem-se revelado eficaz como coadjuvante
para o tratamento desta patologia. Tm-se desenvolvido estudos no sentido de desenvolver
uma terapia gentica para esta condio. Uma das abordagens possveis envolve a utilizao
de um adenovrus modificado no qual o gene para a protena deficiente foi inserido. O vrus
seria, ento introduzido no aparelho respiratrio (ex.: por inalao de um aerossol) de modo
que o epitlio pulmonar passaria a expressar o gene e a produzir a protenas localmente. A
deficincia em o1-antiproteinase est tambm implicada num tipo de patologia heptica.
Nesta condio, as molculas de PiZ sofrem agregao nas cisternas do retculo
endoplasmtico rugoso dos hepatocitos. Estudos recentes revelaram que esta agregao
devida polimerizao das molculas de o1-antiproteinase (que apresentam uma
conformao anormal) atravs do estabelecimento de interaces fortes entre uma ansa de
uma protena e uma estrutura |- pregueada de outra protenas. Por um mecanismo ainda no
determinado, desenvolve-se uma hepatite, que, em 10% dos homozigticos culmina com
uma massiva acumulao de colagnio no parnquima heptico, resultando em fibrose
(cirrose). Tem-se tentado desenvolver igualmente uma teraputica gentica para esta
condio, mas actualmente, o nico meio de tratamento eficaz o transplante heptico.
As hiper-lipoproteinmias familiares - anlise dos dfices de receptores de LDL
Quadro 10: Hiperlipoproteinmias.
AR, autosomal recessive; AD, autosomal dominant; VLDL, very low density lipoprotein;
CHD, coronary heart disease; PVD, peripheral vascular disease; LDL, low-density lipoprotein.
Sebenta de Gentica
FCML
116
Quadro 11: Classificao das hiperlipoproteinmias segundo Frederickson
Doenas peroxissomais:
S S n nd dr ro om me e d de e Z Ze el ll lw we eg ge er r
uma doena autossmica recessiva caracterizada pela inexistncia de peroxissomas em
todos os tecidos. Devido incapacidade de oxidar os cidos gordos de cadeia longa, estes
acumulam-se nos tecidos (principalmente o cido pipeclico e cidos dicarboxlicos),
nomeadamente no tecido cerebral, onde causam graves leses. No plasma e fibroblastos
assiste-se a um aumento de cidos gordos, com diminuio de DHAP-AT nas plaquetas e
fibroblastos. As manifestaes clnicas consistem em hipotonia, hepatomeglia, facies
caracterstico (fissuras e aparncia dismrfica), disfunes neuromotoras. A esperana mdia
de vida reduzida (no mximo 6 anos).
A Ad dr re en no ol le eu uc co od di is st tr ro of fi ia a
uma doena recessiva, relacionada com o cromossoma X. A carncia de enzimas
oxidativas peroxissomais leva a uma incapacidade de degradao dos c. gordos de cadeia
longa, com consequente aumento das suas concentraes plasmticas. Manifesta-se por
uma distrofia do S.N.C. e da suprarenal. O doente sofre de deteriorao mental, que progride
para demncia, afasia, apraxia, disartria e perda de viso.
D Do oe en n a a d de e R Re ef fs su um m - - d da a m mu ut ta a o o p po on nt tu ua al l a ao o d d f fi ic ce e d de e s si in na ai is s d de e t to op po og g n ne es se e
Refsum disease is an autosomal recessive inborn error of lipid metabolism classically
characterized by a tetrad of clinical abnormalities: retinitis pigmentosa, peripheral neuropathy,
cerebellar ataxia, and elevated protein levels in the cerebrospinal fluid (CSF) without an
increase in the number of cells. However, not all patients show all these features. All patients
have accumulation of an unusual branched-chain fatty acid, phytanic acid, in blood and
tissues. The major route for the removal of phytanic acid, which accumulates in Refsum
disease, is via o-oxidation. Mutations in the hydroxylase gene involved in the o-oxidation
result in Refsum disease. Other variable features include cardiac dysfunction, nerve deafness,
ichthyosis, and multiple epiphyseal dysplasia. Increased levels of phytanic acid can also be
Sebenta de Gentica
FCML
117
found in peroxisomal disorders, including Zellweger syndrome and infantile Refsum
syndrome. Most matrix proteins contain at their C-terminus a tripeptide that acts as a
topogenic signal. This peroxisome targeting signal (PTS1), often Ser-Lys-Leu or some variant,
is recognized by a cytosolic binding protein, peroxin 5 (Pex5p). A few other peroxisomal
matrix proteins contain an N-terminal targeting sequence (PTS2) that interacts with Pex7p.
Defects in Pex5p and other proteins involved in the import machinery of PTS1 proteins result
in humans in Zellweger syndrome (ZS) or similar fatal disorders, called peroxisome
biogenesis disorders such as Refsum. At the cellular level, PBDs are characterized by the
absence of morphologically normal peroxisomes. At the biochemical level, the abnormalities
observed in such patients reflect the malfunctioning of all known peroxisomal pathways. The
absence of Pex7p is linked to another severe disorder, rhizomelic chondrodysplasia punctata
(RCDP).
4. Mosaicismo da expresso ligada ao cromossoma X_____________
Mecanismos de inactivao do cromossoma X
X chromosome inactivation is a process that occurs in all mammals, resulting in selective
inactivation of alleles on one of the two X chromosomes in females. It provides a mechanism
of dosage compensation which overcomes sex differences in the expected ratio of autosomal
gene dosage to X chromosome gene dosage (which is 2:1 in males but 1:1 in females). Males
with a single X chromosome are constitutionally hemizygous for X chromosome genes, but
females become functionally hemizygous by inactivating one of the parental X chromosome
alleles. Not all genes on the X chromosome are subject to inactivation; genes which escape
X-inactivation include ones where there is a functional homolog on the Y chromosome, and
some genes where gene dosage does not seem to be important. In rare individuals with an
abnormal number of X chromosomes (45,X; 47,XXX; 47, XXY, etc), a single X chromosome
remains active no matter how many are present. By contrast, in triploid individuals either one
or two X chromosomes remain active and in tetraploids two X chromosomes remain active.
Thus, there must be some kind of counting mechanism to ensure that one X chromosome
remains active for every two sets of autosomes. In mammals, both X chromosomes are active
in the early female embryo. X-inactivation occurs at an early stage in development, being
initiated at the late blastula stage in mice, and most likely also in humans. In each cell that will
give rise to the female fetus, one of the two parental X chromosomes is randomly inactivated
(note that trophoblast cells are an exception; the paternal X chromosome is preferentially
inactivated, which is a classical example of tissue-restricted imprinting). After the paternal or
maternal X chromosome is inactivated in a cell, the same X chromosome usually remains
Sebenta de Gentica
FCML
118
inactive in all progeny cells, that is the X chromosome inactivation pattern is clonally inherited.
This means that female mammals are mosaics, comprising mixtures of cell lines in which the
paternal X is inactivated and cell lines where the maternally inherited X is inactivated. In
addition to X chromosome inactivation in female somatic cells, the X chromosome is known to
be inactivated transiently during gametogenesis in both males and females.
Mecanismo de inactivao do cromossoma X: the process of X chromosome inactivation is
complex, and distinct molecular mechanisms are involved in initiation of inactivation and
maintenance of the inactivation. The X-inactivation center (Xic), which in humans is located at
Xq13, controls the initiation and propagation of X-inactivation. At this centre, the XIST gene
(called Xist in rodents) responsvel pela inactivao do X, no sendo traduzido em protena
mas actuando como RNA inactivador. Xist produz um ncRNA spliced e poliadenilado de
localizao nuclear exclusiva, sujeito a splicing alternativo. um gene de expresso
monoallica. A regulao da expresso do xist complexa e depende de imprinting.
Although the Xist gene is essential for Xic function, Xist alone is not sufficient. The X-
controlling element (Xce) affects the choice of which X chromosome remains active, and is
distinct from Xist, being located 3 to it. In addition, deletion of a 65-kb region 3 to Xist
produces an effect which suggests that elements involved in the counting mechanism lie distal
and 3 to Xist. Recently, another gene has been identified as being transcribed from the
opposite strand to that which is used for transcribing the Xist gene. Because the transcription
unit of the new gene completely overlaps the Xist gene, and is in the reverse orientation, it has
been named Tsix gene. This has given rise to the idea that Xist may be regulated by the Tsix
gene.
Hemoflia e sndrome de Lesh-Nyan
Hemoflia: a hemofilia uma doena recessiva condicionada por um gene localizado no brao
longo do cromossoma X, que provoca um distrbio da coagulao sangunea por deficincia
do factor VIII (na hemofilia A) e do factor IX (na hemofilia B - tambm designada por doena
de Christmas) devido a alteraes disfuncionais dos genes que codificam estes factores. A
sua incidncia de cerca de 1 em 10,000 homens; as mulheres portadoras raramente
apresentam sinais clnicos da doena. O gene para o factor VIII humano foi j sequenciado e
trata-se de um dos genes de maiores dimenses sequenciados at hoje. No de espantar,
portanto, que 4% dos casos, surjam por neomutao. So mltiplas as alteraes detectadas
em indivduos que apresentam diminuio da actividade do factor VIII e incluem deleces
parciais do gene e mutaes pontuais que determinam a terminao prematura da sntese do
polipptido (protenas truncada). O gene para o factor IX foi tambm clonado e sequenciado e
Sebenta de Gentica
FCML
119
foram descobertas cerca de 300 mutaes deste gene, envolvendo, a sua enorme maioria,
mutaes pontuais e algumas deleces parciais. A deficincia de componentes da cascata
de coagulao resulta na formao de uma espcie defeituosa de fibrina, pelo que a
capacidade de coagulao do sangue do indivduo fica comprometida. As manifestaes
clnicas so praticamente iguais nos dois tipos de hemofilia e incluem: distrbios da
coagulao, caracterizados por tempo de hemorragia prolongado; tendncia permanente
para hemorragias espontneas ou traumticas; fcil aparecimento de equimoses;
hemorragias internas dos msculos e articulaes.
Sndrome de Lesch-Nyhan: doena recessiva, ligada ao X. Caracteriza-se pela presena de
uma variante disfuncional da enzima hipoxantina-guanina-fosforribosil-transferase (HGPRT),
que uma enzima chave no metabolismo da purinas (via de salvage das purinas). A sua
actividade cataltica consiste na transformao da guanina e da hipoxantina nos seus
respectivos nucletidos (GMP e IMP), reaces estas que so dependentes da molcula 5-
fosforribosil-1-pirofosfato (PRPP). Quando a actividade desta enzima deficiente, ocorre uma
acumulao de PRPP o qual, por sua vez, intermedirio da via biossinttica das purinas.
Como a sua concentrao intracitoplasmtica o principal parmetro regulador da actividade
desta via, o resultado uma superproduo de purinas e a consequente hiperuricmia.
Apenas os indivduos do sexo masculino so normalmente afectados. Os heterozigticos
podem ser detectados, devido ao facto de apresentarem nveis de actividade enzimtica
inferiores ao normal, mas no so afectados. Pode assistir-se, por vezes, a um mosaicismo
de duas populaes de clulas, uma completamente normal e outra deficiente. Os pacientes
sem nenhuma actividade residual da HGPRT apresentam um fentipo intenso caracterizado
por: paralisia cerebral; atraso mental; hiperreflexia; comportamento compulsivo de auto-
mutilao (morde os lbios e os dedos); sensibilidade perifrica alterada; hiperuricmia; gota
(inflamao recorrente e incapacitante da primeira articulao metatrsico-falngica).
A distrofina e a distrofia muscular de Duchenne
A distrofia muscular de Duchenne uma doena recessiva ligada ao X (brao curto), que
consiste numa patologia muscular atingindo, principalmente, a cintura plvica. Afecta crianas
e jovens, confinando-os a uma cadeira de rodas por volta dos 10 anos. caracterizada pela
ausncia ou deficincia da protena distrofina, que se liga membrana celular na clula
muscular e essencial para transduo da fora gerada pelas miofibrilhas musculares para a
matriz extracelular (membrana basal da clula muscular). Com efeito, a distrofina liga-se, por
um lado aos filamentos de actina de cada sarcmero e, por outro a um complexo
glocoproteico transmembranar, que, por sua vez est ancorado a um componente da
membrana basal designado por merosina. Na ausncia desta protena, a integridade da fibra
Sebenta de Gentica
FCML
120
muscular esqueltica fica comprometida, na medida em que h maior probabilidade de
danificao da fibra em cada contraco muscular, possivelmente devido a um aumento da
permeabilidade para o Ca
2+
, entrando este com maior intensidade para a clula, o que
provoca a activao das proteases celulares que destroem a clula. O defeito bsico
encontra-se no gene estrutural da protena distrofina. O gene est situado no cromossoma X
(Xp21), que possui uma alta taxa de mutao. Em cerca de 1/3 dos pacientes o defeito foi
provocado por neomutao e os restantes 2/3 tm mes portadoras. As manifestaes
clnicas consistem em: fraqueza muscular progressiva; pseudo-hipertrofia dos msculos; sinal
de Gowers (a criana ergue-se "trepando por si mesmo"). Afecta crianas jovens do sexo
masculino e progride rapidamente, conduzindo morte at terceira dcada. A letalidade
precoce impede transmisso por homens afectados. Mutaes allicas no mesmo gene
causam um distrbio mais leve, a distrofia miotnica de Becker. Esta deve-se a uma deleco
parcial do gene para a distrofina, havendo uma menor sntese de mRNA para a distrofina,
baixando os nveis desta, o que tem consequncias ao nvel da contraco e relaxamento
muscular. Assiste-se a uma progressiva e normalmente fatal, fraqueza muscular.
Deficincia em glucose-6-fosfato desidrogenase
O dfice de G6PD provavelmente a mutao clinicamente significativa mais frequente.
Calcula-se que haja mais de 400 milhes de pessoas afectadas. Portugal um pas de baixa
prevalncia de dfice de G6PD, sendo esta estimada em 0,51%. A descoberta desta mutao
ocorreu nos anos 50, quando se procedeu ao estudo do fenmeno hemoltico decorrente da
administrao do anti-malrico Primaquina. Encontrada inicialmente em frica, foi poste-
riormente observada em todo o mundo. Nos ltimos 25 anos foram encontradas cerca de 400
variantes. Embora a maioria dos indivduos afectados seja assintomtica, o risco de ictercia
neonatal e de anemia hemoltica grave aps a toma de certos frmacos, durante infeces ou
aps a ingesto de favas, real e bem conhecido. H tambm uma forma grave e rara de
anemia hemoltica no esferoctica associada a uma forma de deficincia de G6PD. A G6PD
um enzima que catalisa o primeiro passo na via das pentoses fosfato transformando a
glicose-6-fosfato (G6P) em 6- fosfogluconato com reduo do NADP a NADPH. expressa
em todos os tecidos, embora a sua deficincia apenas se faa sentir nos eritrocitos
afectados. A sua funo principal no eritrocito a obteno de NADPH que mantm o
glutatio na forma reduzida, protegendo o mesmo dos efeitos oxidativos dos radicais livres de
oxignio. O gene localiza-se na regio telomrica do brao longo do cromossoma X na banda
Xq28, pelo que os indivduos afectados so, predominantemente, do sexo masculino. Este
gene compreende 13 exes ocupando perto de 20 Kb. Quase todas as mutaes so simples
substituies de uma nica base levando subsequente substituio de um aminocido.
Sebenta de Gentica
FCML
121
Apenas uma variante de G6PD devida a uma deleco (G6PD Gergia), o que leva a
pensar que deleces neste gene no sejam compatveis com a vida.
Fig. 57: Via da pentose fosfato
5. Hereditariedade no convencional____________________________
Dissomias uniparentais
Quando um descendente produz ambas as cpias de um determinado cromossoma derivado
de um nico progenitor dizemos que apresenta uma dissomia uniparental. Por sua vez,
quando ambos os genomas se originam no mesmo progenitor diploidia uniparental- nunca
se desenvolvem correctamente. o caso das molas hidatiformes- conceptos anormais com
um caritipo 46,XX de origem exclusivamente paterna. Os teratomas ovarianos resultam da
diploidia uniparental materna e caracterizam-se por tumores ovarianos benignos.
A dissomia uniparental (UPD) afectando um s par de homlogos no diagnosticada se o
seu resultado no anormal (patolgico) mas detectada quando envolve cromossomas que
produzem sndromes caractersticas. A UPD pode ser uma isodissomia, na qual os dois
homlogos so idnticos, ou uma heterodissomia, na qual derivam dos dois homlogos de
um dos progenitores. Supe-se que a causa geral seja uma tentativa de salvamento: um
concepto trissmico que iria morrer ocasionalmente perde um cromossoma por no-disjuno
mittica ou por anafase tardia de uma clula totipotente. A poro euplide dessa clula
forma o embrio, enquanto todas as clulas aneuplides morrem. Se cada um dos trs
exemplares de homlogos tiver uma chance igual de ser perdido, haver uma chance de 2/3
da perda de um cromossoma levar a uma constituio cromossmica normal e de 1/3 de
ocorrer uma dissomia uniparental (materna ou paterna). A isodissomia uniparental pode
surgir por presso de seleco num embrio monossmico, a fim de alcanar a euploidia por
duplicao selectiva do cromossoma monossmico.
Sebenta de Gentica
FCML
122
Imprinting genmico: sndromes de Prader-Willi e de Angelman
Imprinting genmico: a teoria do imprinting genmico diz que as regies reguladoras dos
genes herdados de cada um dos progenitores, so pr-reguladas diferentemente, pelo que
faro sentir a sua influncia. A expresso diferencial do material gentico ao nvel do
cromossoma ou do alelo herdados designa-se de impriting genmico. Essa expresso
diferencial resultado essencialmente do diferente grau de metilizao dos genes herdados
dos progenitores. Constitui uma excepo s leis de Mendel, pois a expresso gnica
autossmica dependente do progenitor do qual foi herdado. Durante a gametognese tanto
os genes que provm da me como do pai so desmetilados. Depois da fecundao, os
genes so metilados conforme os graus de metilao que possuam na me e no pai. O
processo de metilao levado a cabo por metilases. Quanto menos um gene for metilado
maior ser a probabilidade de um gene se vir a expressar (Modelo de Holliday). O imprinting
tambm se pode dar por excluso mtua de expresso: antisense; metilao diferencial de
elementos vizinhos (cis). Actualmente sabe-se que, num nmero considervel de distrbios
genticos, a expresso do fentipo da doena depende da herana paterna ou materna do
fentipo. Desta forma, no indiferente herdar um gene da me ou do pai pois um deles
poder estar regulado para a induo de uma doena. Dois exemplos so: a forma intensa,
de incio precoce da distrofia miotnica dominante, quando o gene mutante herdado da
me e o incio precoce da doena de Huntington, quando o gene mutante herdado do pai.
Verdadeiramente demonstrativo da importncia da origem parental do material gentico na
expresso clnica de um defeito, podemos referir as sndromes de Prader-Willi e de
Angelman.
Sndrome de Prader-Willi: um sndrome dismrfico, relativamente comum, caracterizado
por: obesidade e polifagia; mos e ps pequenos; baixa estatura; hipogonadismo;
retardamento mental varivel; gesticulao espontnea. Em muitos casos, h uma deleco
citogentica envolvendo a parte proximal do brao longo do cromossoma 15 (15q11q13), que
ocorre no cromossoma 15 herdado do pai. Por consequinte, os genomas destes pacientes
possuem informao gentica em 15q11q13, que provm apenas da me. Pode ser devida a:
65-75% so deleces de novo na regio 15q11- q13 paterna; 20-30% so dissomias
uniparentais maternas (possuem um par intacto de cromossomas 15, sendo ambos os
membros do par de cromossomas, herdados maternalmente), 5% so defeitos de imprinting
em 15q11-q13 paterno. Estudos recentes mostraram que a leso se situava nas smallest
regions overlap (SRO), entre D
15
S
9
e D
15
S
11
.
Sebenta de Gentica
FCML
123
Sndrome de Angelman: tambm conhecido por sndrome do "boneco feliz" ,
fenotipicamente, muito diferente da anterior, caracterizando-se por uma deleco da mesma
regio cromossmica, mas no cromossoma 15 herdado da me.
Por consequinte, os
genomas destes pacientes possuem informao gentica em 15q11q13, que provm apenas
do pai. Pode ser devida a: 65-75% so deleces de novo na regio 15q11- q13 materna, 5%
so dissomias uniparentais paternas, 5% so defeitos de imprinting em 15q11-q13 materno.
Como sinais clnicos apresenta: atraso mental grave; microcefalia; convulses; paroxismos
de riso; movimentos voluntrios compulsivos; ECG normal; macroglossia.
Mola Hidatidiforme: a mola hidatidiforme completa diplide, mas todos os cromossomas so
paternos (androgentica). Nesta origem uniparental, o mecanismo mais frequente a
fertilizao de um ovocito anucleado(ou com ncleo inactivo) por um nico espermatozide,
cujo genoma haplide duplica.
Expanso de tripletos como mecanismo de doena e sndromes de expanso de
tripletos
Foram associadas com o fenmeno da expanso de tripletos de nucletidos dois tipos de
doenas: locais frgeis em cromossomas e doenas neurodegenerativas crnicas.
Cromossomas frgeis: expanses CGG/CCG do origem a locais frgeis nos cromossomas,
como os encontrados na sndroma do X frgil. No entanto, a causa da fragilidade associada
s expanses CGG/CCG ainda no foi encontrada.
Doenas neurodegenerativas associadas expanso de tripletos: expanses do tripleto CAG
esto associadas a pelo menos seis doenas neurodegenerativas, existindo muito
provavelmente outras por identificar. A maior parte destas condies so de manifestao
tardia, sendo uma caracterstica proeminente em todas elas a ocorrncia de morte celular. As
mutaes ocorrem nas open reading frames dos genes envolvidos, sendo as expanses
encontradas mais pequenas do que as identificadas nos cromossomas frgeis (na sua
maioria menores que 100 repeties). As mutaes implicam o ganho de novas funes ou a
sua perda pelas protenas envolvidas consoante a patologia e a confirmao da expanso
pode ser por Southern ou aps PCR.
Fig. 58: Locais de expanso de tripletos
5 5 U UT TR R 3 3 U UT TR R
C Co or re ei ia a d de e H Hu un nt ti in ng gt to on n ( (C CA AG G) )
A At ta ax xi ia a d de e F Fr ri ie ed dr re ei ic ch h ( (G GA AA A) )
D Di is st tr ro of fi ia a m mi io ot t n ni ic ca a ( (C CT TG G) )
S S n nd dr ro om me e d do o X X f fr r g gi il l ( (C CG GG G) )
Sebenta de Gentica
FCML
124
X-frgil
Resulta de perda de funo da protena e desregulao epigentica. uma patologia
dominante ligada ao X, com 80% de penetrncia nos homens, e apenas 30% de penetrncia
nas mulheres. Corresponde a uma zona de fragilidade situada na extremidade distal (5 UTR)
do brao longo (Xq27.3). O marcador do ponto frgil uma constrio prxima da
extremidade distal Xq (regio q27 - q29). No local frgil encontra-se a sequncia CCG que vai
ser grandemente amplificada at atingir um nmero de cpias (cromossomas X normais
contm cerca de 40 cpias, cromossomas portadores de 70 a 200 cpias) que produz
instabilidade ou inactivao do gene FMR-1 uma vez que se encontra hipermetilado na
mutao, silenciando o gene (a funo do seu produto proteico ainda no foi totalmente
identificada sendo que possivelmente ser uma protena de ligao ao RNA que pode
interagir com os ribossomas; possivelmente controlando o transporte de mRNA e a traduo
uma vez que a existncia de NES (Nuclear exportation signal) e NLS (nuclear localization
signal) no gene indica que a protena FMRP exibe um shuttle entre o ncleo e o citoplasma;
a FMRP modula tambm a plasticidade sinptica).
Fig. 59: FMRP e a traduo do RNA
O silenciamento do gene FMR1 um exemplo paradigmtico de um efeito epigentico
causado por variao da sequncia (repetio). O stio frgil s visvel em meio de cultura
especfico - pobre em cido flico (Vit. B12) ou timina ou rico num antagonista do cido flico,
o metatiroxato (meio TC 199). Observado em metafase, o cromossoma X apresenta, na
extremidade distal do seu brao longo, uma espcie de satlite (tal como os acrocntricos)
que no mais do que a poro terminal do cromossoma, separado deste por uma zona de
fragilidade. Esta consiste num alongamento anormal da cadeia de DNA (sequncias de
CCG). A causa da fragilidade associada s expanses CGG/CCG ainda no foi encontrada.
No entanto, os locais frgeis at agora encontrados localizam-se prximo de uma ilha CpG,
parecendo haver uma correlao entre a amplificao de repeties CGG/CCG,
Sebenta de Gentica
FCML
125
hipermetilao e a expresso de locais frgeis. possvel que a metilao possa estabilizar
regies de DNA tetraplxico formado por repeties CCG, o que levaria obstruo da
replicao de DNA e da condensao de cromatina, levando ao estabelacimento de um stio
frgil. Por outro lado, a partir de estudos efectuados no FRA16A, foi sugerido que a metilao
seria uma consequncia em vez de uma causa da gnese de locais frgeis, de modo que o
mecanismo envolvido na fragilidade permanece indefinido. Actualmente, pensa-se que o
referido alongamento de DNA se processa ao longo das geraes e, quando esse
alongamento atinge determinado limiar, a doena manifesta-se - antecipao. Deste modo,
um indivduo do sexo masculino pode possuir a alterao e ser fenotipicamente normal. O
quadro clnico desta sndroma consiste em: atraso mental - QI<50 (especialmente no
desenvolvimento da fala); dismorfia cranio - facial; estatura superior mdia da idade;
macroorquidia (testculos volumosos); orelhas grandes e protuberantes com implantao
baixa; queixo proeminente (prognatismo); alteraes comportamentais (psicticos, autistas,
hiperactivos). As mulheres portadoras no so dismrficas, mas 1/ 3 apresenta leve atraso
mental. As descobertas sobre o padro da mutao nas genealogias explicam o Paradoxo
de Sherman (the risk of expressing mental retardation was dependent on the individuals
position in the pedigree, with risk increasing in later generations. The daughter of an
unaffected male carrier was more likely to have affected offsprings than the mother of the
unaffected male carrier was: something had changed on the X chromosome over the two
generations). To explain the unusual inheritance pattern, Sherman hypothesized that the
alleged gene for fragile X syndrome was mutated in a two-step process. They proposed that
the first mutation caused a premutation state that produced no clinical symptoms, and that a
second mutation was required to convert the premutation to a full mutation form that was
associated with the characteristic symptoms of fragile X syndrome. Moreover, conversion from
a premutation to a full mutation was proposed to occur only when the premutation was
transmitted from a female carrier.
Fig 60: The Sherman Paradox.The daughter (III) of an unaffected male carrier (II) is more likely to have affected
offspring than the mother (I) of the male carrier is.
Sebenta de Gentica
FCML
126
A probabilidade de converso da pr-mutao em mutao completa est relacionada com o
tamanho da amplificao da mulher heterozigota. Assim, um nmero de repeties p(CCG)
variando de 50-70pb apresenta probabilidade pequena de transformao da pr-mutao
completa. Pr-mutaes desse tipo (50-70pb), so apresentadas em mes de homens
normais transmissores. Quando, no entanto, o nmero de repeties est acima de 90pb, o
risco se transformao de pr-mutao em mutao completa prximo ou igual a 100%.
Coreia de Huntington e outras doenas de poliglutamina.
Resultam de ganho de funo da protena.
Coreia de Huntington: ver pgina 100
Doenas de poliglutamina: incluem: Coreia de Huntington, atrofia espino-bulbar, ataxia
espinocerebelar 1, 2 e 3 (doena de Machado-Joseph), Ataxia espinocerebelar 6 e 7.
A patognese por protenas com expanso poliglutamina pode envolver vrias mecanismos:
a) Acumulao da protena mutante com formao de incluses insolveis que recruta
componentes ubiquitina-proteassoma.
b) A protena mutante pode ser degradada por proteases, formando fragmentos patognicos.
c) Pode tambm interferir com a transcrio e metabolismo de mRNA; sendo que a
interferncia com a transcrio pode tambm envolver interaco com factores de transcrio
ou a cromatina.
Doena de Machado-Joseph
Doena neurolgica, de manifestao tardia, em que ocorre uma mutao, num determinado
gene trans-episttico (na extremidade do brao longo de um ou dos dois cromossomas do
par 14), que leva no transcrio do gene (hiposttico) por ele regulado. A DMJ
caracterizada por inmeros problemas a nvel motor como o desequilbrio, a descoordenao
de movimentos, a espasticidade (articulao deficiente e consequente rigidez dos membros),
a limitao dos movimentos oculares (sobretudo para cima e, mais tarde, em todas as
direces). Tambm os olhos ficam salientes e h retraco das plpebras, sem que seja
afectada a capacidade intelectual do indivduo. Os primeiros sintomas so quase sempre o
desequilbrio e as alteraes na marcha. The mechanism responsible for the phenotype
differences in MJD is unknown.
Ataxia de Friedreich
Resulta da perda de funo da protina. A Ataxia de Friedreich foi a primeira forma de ataxia
hereditria a ser distinguida de outras formas de ataxia. uma doena autossmica
Sebenta de Gentica
FCML
127
recessiva. Geralmente os primeiros sintomas da ataxia de Friedreich so observados durante
a infncia ou nos primeiros anos da adolescncia, contudo em alguns indivduos o
diagnstico tem sido efetuado to cedo como aos 2 anos ou to tarde como aos 25 anos. Os
primeiros sintomas so usualmente a dificuldade com o equilbrio, coordenao prejudicada
das pernas ou braos (ataxia) e a dificuldade na articulao das palavras (disartria). Uma
pessoa com ataxia de Friedreich pode ter dificuldades de movimentos precisos com os lbios
e a lngua, tornando seu modo de falar mais vagaroso e de difcil compreenso. A mutao
mais comum na ataxia de Friedreich uma expanso dum segmento de DNA conhecido
como repetio GAA no exo 1 do gene da frataxina- estrutura e localizao mitocondrial
que controla a homeostase de ferro (a frataxina actua como um chaperone do io ferroso
que torna o ferro disponvel ferroquelatase de sntese de heme) e a respirao (complexos
I, II e II da mitocndria); e activadora da fosforilao oxidativa. O trinucleotdeo GAA
repetido entre 7 a 22 vezes em pessoas sem ataxia de Friedreich, mas em pessoas com esta
doena, as seqncias de GAA so repetidas 200 a mais de 1000 vezes. Em geral, quanto
maior a repetio GAA mais precoce e mais severa a doena. At agora quase todos os
pacientes estudados com a ataxia tem a repetio GAA extra em ambos os cromossomas 9,
contudo os segmentos da repetio no tem o mesmo comprimento em cada um deles. A
ataxia de Friedreich a primeira doena associada expanso de tripletos cuja repetio se
situa no intro pois as outras doenas associadas expanso de tripletos apresentam a
repetio no exo. a ataxia hereditria mais comum.
Bases moleculares da ataxia da Friedreich:
a) A expanso GAA no intro 1 de FRDA causa nveis de transcrio diminudos de frataxina.
O nmero de repeties correlaciona-se com a severidade do fenotipo.
b) Frataxina uma protena mitocondrial altamente conservada na escala filogentica.
Parece comportar-se como protena de ligao de Fe; protege a sntese de complexos FeS;
participa no metabolismo do heme.
c) Quando os nveis de frataxina so baixos as protenas com complexos Fe-S (complexos, I,
II e II) e aconitase esto reduzidos. A acumulao de Fe na mitocndria leva formao de
espcies activas de O, o que pode levar a menor sntese de complexos FeS que so
sensiveis ao estado oxidativo.
Distrofia miotnica
Resulta de desregulao epigentica. Miopatia autossmica dominante, notria pela sua
expressividade varivel quanto intensidade clnica e onset age (mais precoce quando
herdada maternalmente). Geralmente, as crianas afectadas com a forma congnita
Sebenta de Gentica
FCML
128
descendem de uma me afectada (esta pode ter apenas uma expresso leve da doena e
nem mesmo saber que afectada).
Fig 61: Antecipao na DM
Corresponde distrofia muscular autossmica dominante mais comum, chegando a ter uma
frequncia de 1/1000 em algumas populaes. A doena provoca: contraces musculares
contnuas, mesmo aps ter cessado o estmulo para a contraco, cataratas, atrofia das
gnadas e calvcie frontal. O principal mecanismo patognico na DM a alterao da
regulao do splicing alternativo de pr-mRNAs: o splicing reverte a formas pr-
embrionrias. A expresso inapropriada de isoformas embrionrias de protenas
responsabilizvel pelas principais manifeses de DM. A alterao de splicing de ClC (canal
de cloro especfico do msculo) e do receptor de insulina (IR), correlacionam-se,
respectivamente, com a miotonia e a resistncia insulina observadas em doentes com DM.
As protenas MBNL e CELF esto envolvidas no splicing alternativo.
Fig. 62: MBNL e CELF
Sebenta de Gentica
FCML
129
6. Patologia citogentica_______________________________________
Citogentica clnica
Estudo que relaciona o aspecto microscpico dos cromossomas e o seu comportamento
durante a diviso celular com o respectivo gentipo e fentipo do indivduo.
As aberraes cromossmicas so responsveis por 60 ou mais sndromas identificveis
que, colectivamente, so mais comuns que todos os distrbios monognicos mendelianos.
Os pacientes com aberraes cromossmicas tm, geralmente, fentipos caractersticos,
sendo as anomalias fenotpicas resultantes do desequilbrio gentico que perturba o curso
normal do desenvolvimento. O atraso do desenvolvimento e os aspectos dismrficos so
comuns a todas as aberraes cromossmicas. As aberraes cromossmicas podem ser:
anomalias estruturais (podem originar-se de novo- risco de recorrncia mnimo- ou serem
herdadas- risco de recorrncia entre 5% a 50% dependento do tipo de anomalia e dos
cromossomas envolvidos) ou anomalias numricas (originam-se espontaneamente sendo o
risco de recorrncia mnimo).
Indicaes para o estudo citogentico: atraso de crescimento; atraso mental; malformaes;
ambiguidade sexual; amenorreia; ginecomastia; infertilidade; abortos de repetio; alteraes
cromossmicas na famlia; defeitos de reparao; fragilidade cromossmica; exposio a
agentes clastognicos; neoplasias.
Polimorfismos e abortamentos de repeti o
Em geral, os cariotipos de pessoas normais do mesmo sexo so bastante semelhantes.
Contudo, detectam-se variaes eventuais na morfologia ou colorao dos cromossomas
denominadas poli/heteromorfismos cromossmicos, que reflectem diferenas de quantidade
ou tipo de sequncias de DNA, em determinada regio ao longo do cromossoma.
Cromossomas homlogos normais podem diferir em tamanho, por muitos milhes de pares
de bases, provavelmente como resultado de diferenas na quantidade de sequncias de DNA
heterocromticas. Os exemplos mais comuns envolvem as regies heterocromticas do
brao longo do cromossoma Y e as regies pericentromricas dos cromossomas dos pares 1,
9 e 16. O polimorfismo cromossmico, embora no sendo patolgica para o prprio, pode
condicionar uma interaco inadequada entre o fuso acromtico, o cinetocoro e o centrmero
durante a meiose, de modo que a probabilidade de gerar gmetas dissmicos ou
monossmicos para um dos referidos pares superior normal. Deste modo, como os
embries com aneuploidias dos pares 1, 9 e 16 so inviveis, estes indivduos podem vir a
sofrer abortamentos de repetio, que, no caso de ocorrerem numa fase muito precoce do
Sebenta de Gentica
FCML
130
desenvolvimento, podem ser confundidos com situaes de infertilidade. Outras regies
heterocromticas tambm exibem variaes considerveis, incluindo os braos curtos dos
cromossomas acrocnticos e a regio centromrica de muitos cromossomas.
Metodologias de bandeamento
As clulas, para anlise cromossmica, devem ter a capacidade de se desenvolver e dividir
rapidamente em cultura. As clulas mais acessveis e que satisfazem esta exigncia so
leuccitos, especificamente os linfcitos T. Para preparar uma cultura destas clulas obtm-
se uma amostra de sangue perifrico, em geral por puno venosa e adiciona-se um
anticoagulante- heparina. De seguida, a amostra centrifugada permitindo a sedimentao
dos linfcitos numa camada distinta. Os linfcitos so colocados num meio de cultura com um
agente mitognico, a fito-hemaglutinina. A cultura incubada a uma temperatura de 37C
durante 48 ou 72 horas, conforme se pretenda observar aberraes cromossmicas ou
elaborar um cariotipo, respectivamente. [Caso se pretenda excluir a hiptese da existncia de
stios frgeis em cromossomas necessrio cultivar as clulas sob condies especficas de
crescimento, utilizado um meio de cultura pobre em cido flico]. Acrescenta-se ento uma
soluo muito diluda de colchicina para impedir a concluso da diviso celular, pela
despolimerizao dos microtbulos do fuso acromtico. Por conseguinte, clulas em
metafase acumulam-se em cultura. De segui\da, adiciona-se uma soluo hipotnica que
causa a tumefaco dos linfcitos, bem como a hemlise dos eritrcitos. Aps vrias
lavagens com fixador para remover a hemoglobina extracorpuscular oxidada, de modo a que
a soluo fique lmpida, lisam-se os linfcitos deixando cair uma gota da suspenso numa
lmina e os cromossomas so finalmente fixados, corados com uma de vrias tcnicas,
ficando assim prontos para anlise. Apesar de ideais para anlises clnicas rpidas, as
culturas celulares preparadas a partir de sangue perifrico tm a desvantagem de ter uma
vida curta (3 a 4 dias). Culturas a longo prazo podem ser feitas a partir de bipsias outros
tecidos, como o tecido epitelial, a medula ssea ou fibroblastos da derme. Por outro lado,
quando se pretende excluir a hiptese de mosaicismo, estes so normalmente os tecidos
Sebenta de Gentica
FCML
131
analisados em adio s clulas de sangue perifrico. Os mtodos de colorao
originalmente disponveis para anlise citogentica no permitiam a identificao dos 24 tipos
de cromossomas (22 autossomas, X e Y). Com efeito, podiam apenas ser classificados em
sete grupos designados pelas letras de A a G (cf. Cariotipo), de acordo com o seu
comprimento total e posio do centrmero.
1. Grupo A (pares 1, 2 e 3) - constitudo pelos cromossomas longos; os cromossomas
1 e 3 so metacntricos, o cromossoma 2 submetacntrico.
2. Grupo B (pares 4 e 5) constitudo igualmente por cromossomas longos, sendo
todos submetacntricos, muito semelhantes entre si, mas distinguveis por tcnicas de
bandeamento.
3. Grupo C (pares 6 a 12) - constitudo por cromossomas de tamanho mdio,
submetacntricos, distinguveis por tcnicas de bandeamento.
4. Grupo D (pares 13 a 15) - constitudo por cromossomas de tamanho mdio,
acrocntricos.
5. Grupo E (pares 16 a 18) constitudo por cromossomas curtos, metacntricos (16)
ou submetacntricos.
6. Grupo F (pares 19 e 20) constitudo por cromossomas curtos, metacntricos.
7. Grupo G (pares 21 e 22) constitudo por cromossomas muito curtos, acrocntricos
(com satlites).
Cromossoma X semelhante aos cromossomas do grupo C (particularmente com o
cromossoma 6), de tamanho mdio e submetacntrico.
Cromossoma Y semelhante aos cromossomas do grupo G, mas um pouco maior,
com os cromatdeos menos divergentes e nunca apresentando satlites.
A identificao inequvoca de cada um dos cromossomas do cariotipo humano s se tornou
possvel depois de 1970, com o aparecimento das tcnicas de bandeamento. Os distintos
mtodos marcam os cromossomas de uma forma diferencial, o que se traduz pelo
aparecimento de regies de coloraes distintas (bandas) com disposio transversal ao
longo dos braos, formando um padro caracterstico de cada cromossoma e que
altamente reprodutvel este facto que confere valor diagnstico anlise de um cariotipo.
Os mtodos diferenciais mais utilizados so os das bandas Q, G e R- revelam eucromatina.
Os cromossomas so observveis em metafase, quando se encontram condensados logo o
nmero de bandas menor comparativamente com as tcnicas de alta resoluo (mais
frente).
Bandas Q foram as primeiras a serem obtidas e o seu mtodo baseia-se na
fluorescncia emitida pela quinacrina, sob a aco dos raios ultra-violeta. Este mtodo
Sebenta de Gentica
FCML
132
permite corar cerca de 50% da eucromatina. Os cromossomas coram num padro especfico
de bandas brilhantes (mais ricas em A-T) e bandas escuras (mais ricas em G-C).
Bandas G a tcnica mais utilizada. Os cromossomas so tratados com tripsina
para desnaturar as protenas cromossmicas. De seguida, coram-se os cromossomas pelo
corante Giemsa. Cora cerca de 50% da eucromatina. O padro de bandas semelhante
entre cromossomas homlogos e distinto entre no homlogos. As bandas G claras so ricas
em G-C e as bandas escuras so ricas em T-A. Os cromossomas em metafase apresentam
cerca de 400 bandas G.
Bandas R surgem por aco de solues alcalinas a quente e aps colorao por
mtodos de fluorescncia ou pelo corante Giemsa. As bandas R produemz um padro de
bandas inverso do das bandas G, ou seja a bandas G positivamente coradas, correspondem
bandas R, negativamente coradas. A caracterstica estrutural dos cromossomas que permite
a colorao diferencial de pores de cromossomas por esta tcnica a existncia de
regies mais ricas em A-T (menos resistentes desnaturao devido a estabelecerem
apenas duas pontes de hidrognio entre si) em determinadas pores cromossmicas e
outras regies mais ricas em G-C, com maior estabilidade trmica. Em alguns casos, o
bandeamento R fornece informaes adicionais s obtidas por outros mtodos de
bandeamento. de salientar que grande parte dos genes conhecidos foram mapeados em
regies cromossmicas correspondentes s bandas R. Apesar de o nmero de cromossomas
conhecido ser ainda pequeno, as anomalias cromossmicas que envolvem as bandas R
(possivelmente mais ricas em genes) parecem ter mais impacto clnico do que as que
envolvem as bandas G (cf. Sequncias CpG e Metilao).
As tcnicas de bandas de alta resoluo so tambm um mtodo diferencial que permite
revelar a heterocromatina. Estas permitem um estudo detalhado, feito nos cromossomas em
profase e pro-metafase. Nestes, o nmero de bandas maior por os cromossomas no se
Sebenta de Gentica
FCML
133
encontrarem ainda totalmente compactados. Constitui um mtodo mais preciso para o
diagnstico de anomalias ocorridas em pores extremamente pequenas do cromossoma,
como microdeleces.
Bandas C - so obtidas pelo mesmo processo que as bandas R e representam a
heterocromatina constitutiva com DNA satlite de replicao tardia (presente nos
cromossomas acrocntricos 1,9 e 16 e poro distal do Y). A identificao dos cromossomas
possvel atravs da colorao dos centrmeros. Permitem observar polimorfismos da
heterocromatina ou outras anomalias cromossmicas.
Bandas NOR - so marcadas pela prata amoniacal nos ncleos em interfase, se esto
activas e nos cromossomas em metafase quando o estiveram na interfase precedente. O
componente responsvel pela colorao uma protena cida que se encontra localizada
junto dos NOR- regies dos organizadores nucleolares que se localizam nos braos curtos
dos cromossomas acrocntricos
O estudo feito ao TEM (microscpio electrnico de transmisso) provou que os tratamentos a
altas temperaturas para a obteno das bandas R levam a alteraes drsticas da estrutura
cromossmica fisicamente visveis. Tal no acontece no caso das bandas G. Segundo alguns
autores, os sulcos que surgem nos cromossomas com bandas G observados ao TEM,
representam essas mesmas bandas.
Sebenta de Gentica
FCML
134
Tcnicas citogenticas moleculares:
Mtodos FISH (fluorescent in situ hybridization) e anlise cromossmica
O princpio deste mtodo a hibridizao de uma sequncia conhecida de DNA ou
cromossmica marcada com um corante fluorescente com a sua sequncia homloga numa
preparao cromossmica convencional seguida da visualizao da sequncia alvo por
tcnicas de fluorescncia, muitas vezes em cores distintas. As etapas de preparao so as
seguintes:
- Preparao e marcao de sondas de DNA com fluorocromos
- Preparao de cromossomas em metfase ou ncleos em interfase ou DNA microarrays
(alta resoluo a nvel do nucletido nico)
- Desnaturao do DNA
- Hibridizao in situ
- Deteco da sonda fluorescente, geralmente por mtodos de microscopia de
fluorescncia
Tem, geralmente, uma elevada sensibilidade e poder de resoluo, sendo possvel a
visualizao de cromossomas inteiros, atravs da utilizao de sondas compostas. Por outro
lado, mesmo alvos de tamanho to diminuto como 500 pb num cromossoma em metfase,
podem ser identificados.Permite o estudo de uma regio cromossmica, a sua representao
numrica e o seu envolvimento em translocaes em qualquer etapa do ciclo celular.
Aplicaes da tcnica:
- Deteco de alteraes numricas dos cromossomas
- Deteco de alteraes estruturais dos cromossomas, e identificao da origem de
cromossomas supra-numerrios e de vrias translocaes
- Deteco de alteraes e aberraes cromossmicas em ncleos interfsicos (o avano
mais importante permitido por esta tcnica)
- Mapeamento gentico
- Estudos filogenticos (evoluo das espcies)
- Deteco de microdeleces
- Estudo facilitado de danos cromossmicos devidos a agentes mutagnicos
No entanto, ainda no substitui adequadamente a citogentica clssica porque:
- S os mtodos clssicos permitem a anlise da totalidade do caritipo em metfase
preparado segundo um nico procedimento;
- Na tecnologia FISH o investigador necessita de uma seleco e orientao prvia da
regio a estudar.
Sebenta de Gentica
FCML
135
- O mtodo FISH ainda no permite a distino entre algumas das regies cromossmicas.
Rx-Fish: Cross Species Color Banding
- Permite identificar alteraes do nmero e estrutura dos cromossomas, visualizando e
analizando cromossomas bandeados numa gama de cores.
- Permite identificar alteraes intracromossmicas e intercromossmicas
- As sondas de DNA so derivadas de espcies filogenticamente prximas da humana
(ex: gibo), so marcadas com fluorocromos e hibridizadas com amostras do genoma
humano
- Baseia-se no facto de que sequncias no repetitivas de DNA humano e de gibo
partilham entre si regies de elevada homologia, embora organizadas da modo diferente
- Assim, uma sonda de DNA construda a partir de gibo pode hibridar com seces no
contnuas de um cromossoma humano e/ou diferentes regies de mais do que um
cromossoma
- A observao feita com microscpio de fluorescncia acoplado a um sistema
computadorizado de anlise de metafases
Multiplex Fish (M-Fish)
- Utiliza vrias combinaes de marcadores fluorescentes, as sondas marcadas so
hibridadas com cromossomas em metafase e assim se obtm cada cromossoma de uma cor
diferente.
- So necessrios 5 fluorocromos para diferenciar os 24 cromossomas humanos
Fibre Fish
- utiliza preparaes de filamentos de cromatina soltos (sem histonas)
- permite grande resoluo
Comparative Genomic Hybridization- CGH
- Envolve a hibridao competitiva de DNA da amostra e DNA de referncia em metfases
- Pode utilizar ncleos em interfase
- No requere nenhum conhecimento prvio da alterao cromossmica envolvida.
- No identifica rearranjos cromossmicos que no envolva perda ou ganho de material
gentico (translocaes equilibradas, inverses, etc), apenas aneuploidias, por exemplo,
amplificao ou deleco de DNA da amostra em relao ao DNA de referncia.
Sebenta de Gentica
FCML
136
Fig. reas de ganho (verde) e perda (vermelho) de DNA tumoral so reveladas quando a razo de DNA tumoral:
DNA controlo mais que 1.5 e menos que 0.5 respectivamente
Aneuploidias de expresso fenotpica patolgica- as trissomias 13, 18 e 21
Aneuploidia: aberrao cromossmica por alterao do nmero de cromossomas em que um
ou mais cromossomas individuais esto excedentes ou ausentes dum conjunto euplide
(euploidia significa possuir conjuntos cromossmicos completos- n, 2n, 3n, etc.).
As causas da aneuploidia ainda no foram completamente esclarecidas mas sabe-se que o
mecanismo cromossmico mais comum a no disjuno meitica.
- A no disjuno na 1 diviso meitica (separao de cromossomas homlogos), origina:
2 gmetas com um cromossoma extra + 2 gmetas com um cromossoma ausente
- A no disjuno na 2 diviso meitica (separao de cromtides irms), origina:
2 gmetas normais + 1 gmeta com um cromossoma ausente + 1 gmeta com um
cromossoma extra
A no disjuno pode tambm ocorrer numa diviso mittica aps a formao do zigoto.
Caso isto acontea, numa diviso por clivagem inicial pode sobrevir mosaicismo
cromossmico, clinicamente significativo.
Outros conceitos importantes:
- poliploidia: sendo a mais comum nas gestaes humanas a triploidia em que dois
espermatozides fertilizam o mesmo vulo ou um dos gmetas diplide.
Sebenta de Gentica
FCML
137
Fig. 62: About two-thirds of human triploids arise by fertilization of a single egg by two sperm (A). Other causes are
a diploid egg (B) or sperm (C). Most human triploids abort spontaneously; very rarely they survive to term, but not
beyond.
Trissomia 13: Sndrome de Patau
Distrbio grave e letal em 50% dos nados-vivos durante o primeiro ms. A esperana mdia
de vida de 130 dias. Como a maioria das outras trissomias, associa-se idade materna
avanada (a distribuio bimodal, possuindo dois picos - um aos 25 anos e outro aos 38
anos). O cromossoma extra provm, geralmente, da no disjuno na meiose I materna (47,
XX ou XY, +13). Cerca de 20% resultam de uma translocao no equilibrada entre os
cromossomas 13 e 14 t (13q, 14q).
Trissomia 18: Sndrome de Edwards
95% resultam em abortos espontneos e diminui de tal modo a sobrevivncia ps-natal, que
esta raramente vai para alm dos 2 meses de idade (no mximo, 10 meses). 80% dos
pacientes so do sexo feminino devido, possivelmente, sua sobrevivncia preferencial. A
causa mais frequente a no disjuno dos cromossomas (47, XX ou XY, +18), podendo
ocorrer mosaicismo em 10% dos casos com manifestaes mais suaves e aumento da
sobrevivncia (mes na faixa etria normal).
Trissomia 21: Sndrome de Down
A incidncia da trissomia 21 de 1 em cada 800 recm-nascidos. O risco varia com a idade
da me, aumentando concomitantemente com a idade materna. Em 95% dos pacientes, a
sndrome de Down envolve trissomia do cromossoma 21, resultante da no disjuno
meitica do par de cromossomas 21, predominantemente na meiose I materna (95%),
podendo igualmente ocorrer na meiose I paterna. Cerca de 4% dos pacientes apresentam 46
cromossomas, um dos quais uma translocao robertsoniana entre o cromossoma 21 (21q)
e outro dos demais cromossomas acrocntricos (geralmente o cromossoma 14 ou 22). O
cromossoma com translocao substitui um dos acrocntricos normais e, portanto, o cariotipo
de um paciente com uma translocao robertsoniana entre os cromossomas 14 e 21 46, XX
ou 46,XY, -14, + t (14q21q), logo, o paciente trissmico para 21q (ver pgina 143). Cerca de
1% dos pacientes com Sndroma de Down tem mosaicismo gonadal, geralmente, com
populaes celulares com um cariotipo normal ou trissomia do par 21, isto , clulas com 46
e com 47 cromossomas. Nestes casos, as manifestaes so mais ligeiras e o atraso mental
menor.
Regio crtica em 21q22 (Down Syndrome critical region- DSCR)
DSCR1- A sobre-expresso de DSCR1 inibe a transcrio gnica calcineurina-dependente.
Sebenta de Gentica
FCML
138
DSCR2- Protena rica em leucina (expresso no crebro, colon, leucocitos, mama e
testculo).
DSCR3- Homlogo de um gene de ratinho essencial na embriognese (crebro, pulmo,
fgado e rim fetais e de adulto).
DYRK- Serina/treonina protena cinase; pode estar envolvida no desenvolvimento cerebral
normal; defeitos de aprendizagem dose-sensveis.
DSCAM- Molcula de adeso celular, expressa no desenvolvimento cardaco, dose-sensvel.
SOD1- Superoxide dismutase gene.
APP- amyloid beta A4 precursor protein. A quase totalidade dos doentes com sndrome de
Down que vivem para alm dos 40 anos desenvolve manifestaes da doena de Alzheimer.
Este facto poder dever-se ao efeito da dosagem gnica devida trissomia para o gene APP
que codifica a protena precursora da amilide e est localizada no cromossoma 21. A
mutao deste gene est associada a casos familiares de doena de Alzheimer.
Risco de recorrncia da sndrome de Down:
- Trissomia livre: menos de 1% de recorrncia at aos 35 anos
- Translocao: risco acrescido- 1/3 de probabilidade. Se a translocao for de origem
materna o risco ainda maior
- Mosaicismo gonadal: risco acrescido
Sndrome de Turner
Achados citogenticos:
45, X0 (monossomia do X em 60% dos casos)
46, X, i(Xq) ou i(Xp) (isocromossoma X em 5-10% dos casos)
Mosaicismo, anel, deleces, etc.
Dados clnicos:
- Pequenas anomalias morfolgicas: ex pterygium colli
- Taxa de crescimento lenta. Por vezes dificuldades de aprendizagem
- Puberdade assinalada apenas por pilosidade escassa. Amenorreia primria.
- Comprometimento do desenvolvimento sseo com o estabelecimento de estatura mdia de
1,40 m (+90%) gene SHOX. O gene SHOX (Xpter-p22.32) de short stature homeobox-
containing gene um gene de transcrio com homeodomnio (homeodomnio o motivo de
ligao ao DNA (helix-turn-helix de 3 -hlices)). Muitos dos genes Hox esto envolvidos na
Sebenta de Gentica
FCML
139
regulao do desenvolvimento, diferenciao e organognese e tm expresso espacial e
temporal reguladas. Os genes Hox so codificados por sequncias com homeobox. O gene
SHOX exibe haploinsuficincia; i.e. situao em que, ou por hemizigotia devida a deleco,
ou por heterozigotia devida a mutao, o nico alelo presente no suficiente para garantir a
funo normal do gene o que no compensado pelo gene SHOX2 no cromossoma 3.
Deleces e Microdeleces
So rearranjos estruturais que resultam de quebra cromossmica seguida de reconstituio
numa combinao anormal. Podem ser deleces de novo; translocaes em no-equilbrio
ou mendelianas. A troca de cromossomas ocorre espontaneamente numa baixa frequncia e
tambm pode ser induzida por agentes causadores de quebras (clastognicos), como a
radiao ionizante, algumas infeces virais e muitas substncias qumicas. Tal como as
alteraes numricas, os rearranjos estruturais podem estar presentes em todas as clulas
de um indviduo ou sob a forma de mosaico. As deleces e microdeleces so rearranjos
no equilibrados uma vez que h perda/ausncia de parte da informao gentica.
Sndrome do grito de gato e variabilidade das extenses de deleco de 5p
Resulta da deleco parcial do brao curto (p) do cromossoma 5 que ocorre, geralmente, na
zona 5p14p15p. Na maioria dos casos so espordicos, apesar de 10 a 15% serem
descendentes de portadores de translocaes. Raramente se observam mosaicos,
translocaes de novo no equilibradas ou um cromossoma em anel.
A sndroma do "cri-du-chat" (assim chamada pois os doentes apresentam um choro
caracterstico que se assemelha ao miar de um gato) a afeco mais comum associada a
alteraes estruturais. Tendo em conta que os cromossomas so estruturas frgeis podem
sofrer quebras espontneas, ou induzidas por determinados agentes clastognicos. Os
cromossomas que sofram este tipo de leso, tendem a ser reconstitudos atravs da fuso
das extremidades quebradas. Na maior parte das vezes, a reunio correcta e a leso
resolvida, no entanto, podem aparecer cromossomas com anomalias estruturais.
Sndrome de DiGeorge
(ver sndrome dos genes contnuos, pg. 144)
Microdeleco do cromossoma Y e infertilidade
Microdeleces do cromossoma (YCM) faz parte da famlia de doenas genticas causadas
por perdas de genes no cromossoma Y. A maior parte dos homens com YCM vive uma vida
normal e no apresenta qualquer tipo de sintomas. No entanto, h uma relao entre YCM e
Sebenta de Gentica
FCML
140
infertilidade (20% dos homens com reduzida produo de esperma foram identificados como
tendo YCM). A reduzida produo de esperma denominada oligozospermia sendo a
azospermia (mais frequente na YCM) a ausncia total de esperma. No cromossoma Y foram
mapeados trs locais denominados factores de azospermia (AZFa, b e c) onde se encontram
diferentes de genes que esto envolvidos na espermatognese (DAZ, RBMY, USP9Y e
DBY). Deleces nestas regies removem um ou mais destes genes levando uma
testiculopatia severa que provoca infertilidade. No h relao clara e linear entre o tamanho
e a localizao das deleces e o fentipo testicular no entanto, bvio que quanto maior a
rea delectada maior o dano testicular causado. Um problema que advm do YCM o facto
de que os doentes podem ser dadores de esperma (uma vez que muitos manifestam apenas
uma reduo da quantidade de esperma) levantando uma sria de problemas ticos e
clnicos uma vez que caso o dador doente seja utilizado, vai, invariavelmente, passar este
dfice gentico ao descendende masculino.
Deleces em 1p e 19q em oligodendrogliomas, sig. prognstico e teraputico
Oligodendrogliomas are a type of glioma that are believed to originate from the
oligodendrocytes of the brain or from a glial precursor cell. They occur primarily in adults
(9.4% of all primary brain and central nervous system tumors) but are also found in children
(4% of all primary brain tumors). The average age at diagnosis is 35 years. By far, the most
common structural deformity found is co-deletion of chromosomal arms 1p and 19q. The high
frequency of co-deletion is a striking feature of this glial tumour and is considered as a
"genetic signature" of oligodendroglioma. Allelic losses on 1p and 19q, either separately or
combined, are more common in classic oligodendrogliomas than in either astrocytomas or
oligoastrocytomas. In one study, classic oligodendrogliomas showed 1p loss in 35 of 42 (83%)
cases, 19q loss in 28 of 39 (72%), and these were combined in 27 of 39 (69%) cases; there
was no significant difference in 1p/19q loss of heterozygosity status between low-grade and
anaplastic oligodendrogliomas. 1p/19q co-deletion has been correlated with both
chemosensitivity and improved prognosis in oligodendrogliomas. The gene products lost as a
consequence of this codeletion may include mediators of resistance to genotoxic therapies.
Alternatively, 1p/19q loss might be an early oncogenic lesion promoting the formation of glial
neoplasms, which retain high sensitivity to genotoxic stress. Most larger cancer treatment
centers routinely check for the deletion of 1p/19q as part of the pathology report for
oligodendrogliomas. The status of the 1p/19q loci can be detected by FISH or virtual
karyotyping. Virtual karyotyping has the advantage of assessing the entire genome in one
assay, as well as the 1p/19q loci. This allows assessment of other key loci in glial tumors,
such as EGFR and TP53 copy number status. Whereas the prognostic relevance of 1p and
Sebenta de Gentica
FCML
141
19q deletions is well established for anaplastic oligodendrogliomas and mixed
oligoastrocytomas, the prognostic relevance of the deletions for low-grade gliomas is more
controversial. In terms of low-grade gliomas, a recent study also suggests that 1p/19q co-
deletion may be associated with a (1;19)(q10;p10) translocation which, like the combined
1p/19q deletion, is associated with superior overall survival and progression-free survival in
low-grade glioma patients. Oligodendrogliomas show only rarely mutations in the p53 gene,
which is in contrast to other gliomas. Epidermal growth factor receptor amplification and whole
1p/19q codeletion are mutually exclusive and predictive of completely different outcomes, with
EGFR amplification predicting poor prognosis. There is a strong correlation between 1p/19q
codeletion and the expression of proneural genes, suggesting that gliomas with a 1p19q
codeletion represent a subgroup of proneural gliomas.
Molecular analyses of human malignancies began with the observation
that the distribution
of chromosomes had been corrupted in the
nuclei of cancer cells. Methods have since evolved
to expose
the smallest of genetic differences between normal and tumor
cells, and the hope is
that some of these lesions can be targeted
therapeutically. Although specific genetic or
expression patterns
are the basis for the design of the next generation of cancer
therapies, the
chemosensitivity of tumors to existing regimens
has already been associated with certain
genetic alterations in
some human cancers. An example of this was found in anaplastic
oligodendroglioma,
in which tumors showing combined loss of 1p and 19q exhibited
a clinical
behavior distinct from tumors in which 1p and 19q were
intact.Specifically, patients whose
tumors demonstrated
1p/19q loss lived significantly longer and their tumors were
more
sensitive to therapeutic agents. In addition, the clinical diagnosis
of brain tumors is based
primarily on morphological features, and
a goal of molecular diagnosis would be to serve as
an adjunct
to histopathology with respect to classifying subtypes of brain
tumors.
These are
compelling results to use the molecular analysis of chromosomes
1p and 19q to assist in the
histological diagnosis of human gliomas
and clinical management of affected patients. Several
molecular methods
are currently used to detect 1p/19q loss in oligodendrogliomas in
both
clinical and basic research settings. Allelic imbalance, sometimes
referred to as loss of
heterozygosity (LOH), detects reduction
from heterozygosity to homozygosity at 1p and 19q
loci. The routine use of LOH to detect alterations at specific loci
is limited by the requirements
of paired normal tissue from
the same patient and the presence of heterozygosity in the
normal tissue
(an informative locus). Fluorescence in situ hybridization (FISH)
and
comparative genomic hybridization are established methods
to detect copy number, but both
are labor-intensive techniques.
Real-time quantitative polymerase chain reaction (PCR) is
currently
also used to determine copy number at genomic loci but, to date,
only single-copy
sequence-specific probes have been used.
Sebenta de Gentica
FCML
142
Sndrome dos genes contnuos (DiGeorge, Smith-Magenis e Williams)
Sndrome dos genes contnuos:
- microdeleces em determinadas regies de um cromossoma levam a uma doena que
apresenta uma sobreposio de caractersticas de diferentes doenas mendelianas ligadas
ao cromossoma em questo.
- situaes devidas a deleco ou duplicao de mltiplos genes em loci adjacentes. Estes
sndromes caracterizam-se por diversas manifestaes clnicas aparentemente no
relacionadas. Cada um dos genes da regio atingida originar, se mutado ou delectado, um
quadro clnico diverso.
Exemplo do menino BB: sofria de distrofia muscular de Duchenne; doena granulosa crnica
e retinite pigmentar bem como atraso mental. O que se passava com o menino BB que
tinha uma microdeleco em Xp21 que removera um conjunto de genes contnuos logo
manifestava sintomas de diversas doenas (associadas ao genes que foram delectados)
Sndrome de Williams (WLS): pessoas com WLS tm atraso de crescimento, hipercalcmia
na infncia e frequentemente apresentam uma estenose artica supravalvular (SVAS). SVAS
pode ocorrer de forma isolada ou pode ser uma doena autossmica dominante. Esta foi
mapeada no cromossoma 7q11.23 e mostrou ser o resultado da deleco ou disrupo de
alguns genes incluindo o gene da elastina. Muitos outros genes foram encontrados
delectados na WLS embora o seu papel na doena no seja ainda muito claro.
Sndrome Smith-Magenis: um syndrome que afecta 1 em 25 000 indivduos. Resulta duma
deleco duma regio do cromossoma 17 que contm, entre outros genes, o gene RAI1
(17p11.2). Este gene codifica uma protena cuja funo permanence desconhecida mas que,
a falta de expresso desta protena leva aos problemas fsicos, mentais e comportamentais
dos deontes com sndrome Smith-Magenis. um sndrome que no hereditrio mas resulta
de uma alterao gentica na formao dos gametas ou do zigoto. Algumas caractersticas
de pessoas com esta syndrome incluem problemas de comportamento (agresso, auto-
mutilao, auto-abraos repetitivos, lambidelas complusivas nos dedos e folheamento de
livros e revistas compulsivo- comportamento lick and flip).
Sndrome DiGeorge: um sndrome congnito raro cujos sintomas incluem alterao dos
traos faciais, defeitos cardacos e histria de infeco recorrente (um dos genes delectados
neste sndrome necessrio para o bom funcionamento do Timo logo h perda das T-cells.
O syndrome DiGeorge causado pela deleco de uma grande poro do cromossoma 22
resultante de um erro de recombinao na meiose. Um dos genes afectados o TBX1. Este
possui um extenso domnio N terminal de ligao ao DNA (T-box). membro de uma famlia
gnica de ortlogos filogeneticamente conservados com T-box. A T-box do DNA codifica um
Sebenta de Gentica
FCML
143
domnio de ligao ao DNA. Os membros desta famlia gnica parecem reconhecer e ligar-se
todos sequncia de consenso TCACACCT do DNA. Encontram-se exclusivamente no
ncleo e tm actividade de regulao da transcrio estando envolvidos no desenvolvimento
embrionrio. Deleco ou mutao de TBX1 causa alterao da migrao de clulas da crista
neural necessrias ao desenvolvimento dos arcos farngeos.
Cromossomas em anel e isocromossomas
Tanto os cromossomas em anel como os isocromossomas so rearranjos cromossmicos
estruturais no equilibrados.
Isocromossomas: resultam de uma m diviso do centrmero que se faz no sentido
transversal em vez de longitudinal, resultando num cromossoma que duplicao de um
nico brao. Assim, um isocromossoma um cromossoma que tem dois braos
geneticamente iguais. O caso mais frequente um isocromossoma do brao longo do
cromossoma X, i (Xq) em indivduos com a sndrome de Turner (a mulher afectada,
monossmica para os genes do brao curto do X e trissmica para o brao longo).
Fig. 63: Isocromossoma
Cromossomas em anel (ou anel cromossmico): resulta da quebra e perda das extremidades
de um cromossoma (telmeros) com posterior unio dos pontos de quebra- cntricos ou
acntricos, consoante incluem ou no o centrmero (ver fig.64). Os fragmentos contendo os
telmeros so perdidos durante a diviso celular. Como h perda de material gentico, os
indivduos com cromossomas em anel vo assemelhar-se aos que apresentam uma deleco
correspondente. O anel uma estrutura instvel. Pelo fenmeno da troca de cromtides
irms (SCE), que ocorre durante a mitose, a partir do anel cromossmico pode forma-se um
anel dicntrico, porque aps a diviso do centrmero, as duas cromtides ficam unidas pelas
zonas terminais.
Outras anomalias estruturais (dicntricos, acntricos, entre outros)
Sebenta de Gentica
FCML
144
Cromossomas Dicntricos: cromossomas com dois centrmeros
Cromossomas Acntricos: cromossomas sem centrmero
Inverses pericntricas e paracntricas
Inverso: um rearranjo cromossmico estrutural equilibrado que resulta de duas quebras,
no mesmo cromossoma, com rotao de 180 do segmento entre estas e posterior unio dos
pontos de quebra. Conforme envolva ou no o centrmero, dita pericntrica ou paracntrica
(ver fig. 64). Uma inverso isolada no parece conduzir a um fentipo anormal no homem. A
relevncia mdica das inverses para a gerao subsequente e deriva das consequncias
de um crossing-over entre um cromossoma normal e um invertido.
Fig. 64: Resultados estveis possveis de duas quebras num nico cromossoma
Translocaes equilibradas, transmisso e clculo de risco
As translocaes so rearranjos cromossmicos estruturais equilibrados que envolve a troca
de fragmentos cromossmicos entre cromossomas no homlogos. O processo requer
quebra em ambos os cromossomas com um rearranjo anormal.
- Se h troca de fragmentos entre dois cromossomas, a translocao dita recproca
(ver fig. 64 e 65). Esta consiste na troca de material entre dois cromossomas no homlogos
No tm segregao estvel
na mitose e eventualmente
sero perdidos
Sebenta de Gentica
FCML
145
sem que ocorra ganho ou perda de material gentico, pelo que a translocao equilibrada,
conduzindo a um fentipo normal. No entanto, as translocaes recprocas podem ter como
consequncia a formao de gmetas em desequlibrio (alto risco de descendncia anormal).
Isto deve-se formao de tetrarradiais, durante a profase I da meiose, o que conduz a uma
segregao anormal.)
- Translocao Robertsoniana (ou fuso cntrica), um tipo particular de translocao
recproca em que os pontos de quebra esto localizados nos centrmeros de dois
cromossomas acrocntricos ou muito prximo destes (ver fig. 66). Os dois centrmeros esto
presentes mas funcionam como um s (da o termo fuso cntrica) e o cromossoma
estvel. O pequeno frangmento acntrico perdido mas isso no tem consequncias
patolgicas uma vez que o fragmento contm apenas sequncias repetidas de genes de
rRNA que tambm esto presentes noutros cromossomas acrocntricos. A translocao
robertsoniana, embora no tenha consequncias para o indivduo em questo, acarreta
riscos para a sua descendncia. Por exemplo, quando um dos acrocntricos envolvidos o
21, existe uma grande probabilidade do portador originar gmetas com dois cromossomas 21
(fig. 66). Deste modo, existe uma grande risco de uma descendncia com sndrome de Down,
embora o progenitor seja fenotipicamente normal.
Fig. 64: Origem das translocaes
Sebenta de Gentica
FCML
146
Fig. 65: Resultado da meiose num portador de uma translocao recproca balanceada
Fig. 66: Resultado da meiose num portador duma translocao Robertsoniana
Nomenclatura das alteraes cromossmicas estruturais:
Deleco: 46, XY,del(4)(p16.3), 46,XX,del(5)(q13q33)
Inverso: 46,XY,inv(11)(p11p15)
Duplicao: 46,XX, dup(1)(q22q25)
Insero: 46,XY, ins(12;10)(q15;q21.2q22.1)
Anel: 46, XY, r(7)(p22q36)
Sebenta de Gentica
FCML
147
Translocao reciproca: 46,XX, t(2;6)(q35;p21.3)
Translocao robertsoniana: 45, XY,der(14;21)(q10;q10); 46, XX,der(14;21)(q10;q10),+21
Mosaicismo citogentico
O mosaicismo corresponde existncia de mais do que uma linhagem celular geneticamente
diferentes num indivduo, proveniente de um nico zigoto. Assim, um mosaico cromossmico,
tem pelo menos 2 linhagens de clulas com diferentes cariotipos, cujas alteraes podem ser
numricas ou estruturais. Resulta, geralmente, duma no disjuno cromossmica ou perda
de cromossomas por instabilidade do cariotipo numa diviso mittica ps-zigtica inicial. O
mosaicismo pode afectar tecidos da linhagem somtica e/ou da germinativa. As mutaes
ps-zigticas no so apenas frequentes, so inevitveis. Cada um de ns deve ser um
mosaico com inumerveis doenas genticas. Se uma clula do nosso ouvido desenvolver
uma mutao para a fibrose qustica no haver qualquer problema para ns ou nossos
descendentes. S haver risco para o organismo como um todo se uma mutao somtica
resultar no aparecimento de um clone de clulas mutantes com extenso significativa. Isto
pode acontecer de duas maneiras:
- a mutao causa uma proliferao anormal de clulas que normalmente se multiplicariam
pouco- clone de clulas mutantes. Esta a maneira pela qual o cancro pode ocorrer.
- a mutao ocorre no embrio inicial, afectando uma clula que ser a progenitora de uma
parte significativa do organismo total. O indivduo-mosaico pode apresentar sinais da doena.
As mutaes que ocorrem na linhagem germinativa de um progenitor podem causar uma
doena hereditria de novo num filho. Quando um indivduo possui um clone amplo de
clulas germinativas mutantes (mosaicismo gonadal ou germinativo) resultante de uma
mutao ocorrida nos primrdios da formao da linhagem germinativa, um casal normal,
sem histria familiar prvia pode produzir mais de uma criana com a doena dominante
grave. Por exemplo, cerca de 1% dos doentes com sndroma de Down apresentam
moisaicismo e tm uma mistura de tecidos com 46 e 47 cromossomas.
Fig. 67: The genetic change indicated may be a gene mutation, a numerical or structural chromosomal change, or
in the special case of lyonization, X-inactivation.
Sebenta de Gentica
FCML
148
Quimera: em zoologia- animal que tem duas ou mais populaes de clulas geneticamente
distintas que teve origem em diferentes zigotos.
Em seres humanos acontece quando dois vulos fecundados se fundem antes do quarto dia
de gestao misturando as informaes genticas sem que o indivduo sofra grandes
mutaes. Em seres humanos raro; registam-se apenas 40 casos.
Fig. A chimera is derived from two zygotes, which are usually both normal but genetically distinct.
Fragilidade cromossmica e mapa de locais frgeis
Fragilidade cromossmica: constitui um grupo especial de sndromes recessivas raras,
caracterizadas por uma excessiva instabilidade dos cromossomas. Uma deficiente reparao
do DNA pode provocar alteraes ao nvel dos cromossomas ou das cromtides, o que
possibilita a ocorrncia de stios frgeis que se quebram facilmente e que apresentam uma
susceptibilidade elevada a agentes clastognicos. Isto confere ao cromossoma uma
excessiva instabilidade. A sua manifestao fenotpica depende das condies ambientais.
Na deteco de fragilidade cromossmica podemos encontrar diversas aberraes
estruturais nas clulas em estudo lacunas nas cromtides chromatid gap (ctg), quebras nas
cromtides chromatid break (ctb), lacunas nos cromossomas chromossome gap (csg),
quebras cromossmicas chromossome break (csb), dicntricos (dic), tricntricos, acntricos
(ac), trirradiais, quadrirradiais e rearranjos. As sndromes de fragilidade cromossmica e a
incidncia de neoplasias esto fortemente relacionadas visto que os indivduos com tais
sndromas apresentam uma maior frequncia de quebras cromossmicas espontneas
(associadas a mecanismos defeituosos de reparao de DNA). A distino entre os
sndromes de fragilidade cromossmica e as aberraes cromossmicas de origem congnita
baseia-se no facto de que, enquanto na fragilidade cromossmica o cariotipo do indivduo
normal, existindo apenas uma dada percentagem de clulas que revelam essa instabilidade
(no tecido afectado), nas aberraes cromossmicas congnitas o cariotipo anormal.
Da clnica citogentica e anlise molecular do sndrome do X frgil
Sebenta de Gentica
FCML
149
Ver syndrome do X frgil, pgina 126.
7. Patologia Multifactorial______________________________________
Situaes multifactoriais
The further away a character is from the primary gene action, the less likely is it to show a
simple mendelian pedigree pattern. DNA sequence variants are virtually always cleanly
mendelian- which is their major attraction as genetic markers. Protein variants (electrophoretic
mobility or enzyme activity) are usually mendelian but can depend on more than one locus
because of posttranslational modification. The failure or malfunction of a developmental
pathway that results in a birth defect is likely to involve a complex balance of factors. Thus the
common birth defects (cleft palate, congenital dislocation of the hip, congenital heart disease,
etc.) are rarely mendelian. Behavioral traits like IQ test performance or schizophrenia are still
less likely to be mendelian. This does not however mean that they may not be genetic, either
partly or entirely. Nonmendelian characters may depend on two, three or many genetic loci,
with greater or smaller contributions from environmental factors.
Fig. 68: The spectrum of human characters
We use multifactorial here as a catch-all term covering all these possibilities. More specifically,
the genetic determination may involve a small number of loci (oligogenic) or many loci each of
individually small effect (polygenic); or there may be a single major locus with a multifactorial
background. For dichotomous characters the underlying loci are envisaged as susceptibility
genes, while for quantitative characters they are seen as quantitative trait loci (QTLs). Recent
developments have finally brought together the study of mendelian and complex human
phenotypes. Automation is allowing genetic analysis and sequencing on a scale scarcely
imagined ten years ago. This has had two consequences. Most human genes are now
identified, at least as expressed sequence tags (ESTs), so that molecular geneticists are
looking for fresh fields to conquer. At the same time, marker studies can now be done on a
Sebenta de Gentica
FCML
150
scale that is probably large enough to deliver the statistical power needed to detect individual
quantitative trait loci and susceptibility loci. Given the overwhelming preponderance of
nonmendelian conditions in human disease, molecular dissection of complex phenotypes is
widely seen as the next frontier in medical genetics.There are not two sorts of genes,
mendelian genes and polygenes. It is possible however that there are two sorts of mutations.
The sequence variants that confer susceptibility to polygenic disease may not, for the most
part, be the gross mutations seen in mendelian conditions, which typically totally inactivate a
gene. They are likely to include more subtle variants that slightly modify the expression of a
gene. These will exist as common nonpathogenic variants in the healthy population, that
cause problems only when combined with a whole series of similar variants at other loci.
Identificao de genes de susceptibilidade em doenas multifactoriais
As doenas multifactoriais/complexas resultam da interao dos produtos de transcrio de
numerosos genes com mltiplos factores do meio ambiente. A identificao de loci ou de
alelos de susceptibilidade gentica (regies de susceptibilidade individual para o
desenvolvimento de doenas como a hipertenso ou cancro) depende do desenvolvimento
de marcadores polimrficos. Dois tipos de estudos so utilizados na identificao de loci ou
de alelos de susceptibilidade: os estudos de anlise de linkage e os estudos de associao.
Ambos dependem da anlise de polimorfismos genmicos. O contedo informativo dos
marcadores depende do nmero de alelos possveis para cada marcador.
This strategy has already led to the identification of susceptibility genes (involved in the
genesis of the disease) or modifier genes (which may modulate the severity of the disease
once it has developed) involved in heart failure of idiopathic cause (dilated cardiomyopathy) or
of ischaemic origin.
Exemplos de doenas com genes de susceptibilidade:
- Cancro da mama
- Doena de Hirschsprung (doena oligognica clssica com vrios lcus identificados que
contribuem para a susceptibilidade, mas nenhum deles , por si s, necessrio ou suficiente);
- Doena de Alzheimer (factores genticos importantes tanto na forma comum, de incio
tardio, quanto nas formas mendelianas raras, de incio precoce, mas so genes diferentes)
- Diabetes melitus tipo I: a anlise de pares de irmos afectados definiu lcus de
susceptibilidade, e o desiquilbrio de linkage est a ser usado para definir factores genticos.
Estudos de associao de caso-controlo e estudos familiares
Estudos de associao
Sebenta de Gentica
FCML
151
Baseiam-se na deteco da expresso de um determinado marcador fenotpico como
indicativo da presena de um gene herdado em associao e responsvel pela expresso de
um determinado trao ou caracter. Esta abordagem de natureza indirecta. O marcador
associado a um elevado risco para uma doena no o responsvel pela doena. O que se
compara a incidncia de um polimorfismo marcador na populao de indivduos afectados
por uma determinada doena, com a incidncia do mesmo marcador numa populao
controlo. Ambas as populaes tm que ter a mesma origem. Se detectada uma diferena
significativa, esta diferena considerada como indicador de associao positiva ou negativa.
Os estudos de associao tm vantagens sobre os estudos de ligao gnica uma vez que
podem ser realizados em conjuntos de indivduos sem relao de parentesco, seja para os
doentes seja para os controlos, e podem ainda detectar efeitos genticos que os estudos de
ligao gnica no podem detectar. Contudo, dever haver cuidado na comparabilidade dos
grupos de doentes e controlos, considerando que as frequncias allicas esto sujeitas a
grande variao. Por outro lado, a taxa de mutao do alelo estudado e do presumvel lcus
de susceptibilidade deve ser baixa ou conhecida. H ainda vantagem em estudar loci que
codifiquem protenas que possam estar eventualmente relacionadas com a doena.
Nos resultados dos estudos de associao podem-se encontrar duas possibilidades, atravs
do clculo do odds ratio (OR- estimativa do risco relativo nos estudos caso-controlo) e
respectivo intervalo de confiana (IC):
- a frequncia do alelo estudado no difere significativamente entre o grupo de doentes e o
grupo controlo (IC englobando a unidade)
- a frequncia do alelo difere significativamente entre as duas populaes (IC no englobando
a unidade)
Ex:calculo de risco relativo (RR) evidencia numa populao, a associao entre a presena
do alelo polimrfico HLA- B27 e a ocorrncia de espondilartrite anquilosante.
HLA-B27 (+) HLA-B27 (-)
Doentes 210 19
Controlo 42 361
(+) presena do alelo; (-) ausncia do alelo
Clculo do risco relativo: (210/42) / (19/361) = 95. Significa que os portadores do alelo HLA-
B27 tm uma probabilidade 95 vezes maior de desenvolver espondilartrite anquilosnate em
comparao com portadores de outros alelos HLA
Um valor de associao como encontrado neste exemplo excepcional em comparao com
os valores relativamente baixos observados habitualmente para doenas multifactoriais. Os
Sebenta de Gentica
FCML
152
valores baixos podem dever-se ao reduzido contributo do locus estudado comparativamente
com outros loci e com o meio ambiente ou a desiquilibrio de linkage.
O recurso aos grupos sanguneos para estudos de associao foram os primeiros a serem
utilizados mas apresentam limitaes. Os SNPs tambm se encontram ente os marcadores
polimrficos que podem ser usados para estudos de associao.
Estudos Familiares
a) Estudos de gmeos
Monozygotic (MZ) twins are genetically identical clones and will necessarily be concordant
(both the same) for any genetically determined character. This is true regardless of the mode
of inheritance or number of genes involved; the only exceptions are for characters dependent
on postzygotic somatic genetic changes. Dizygotic (DZ) twins share half their genes on
average, the same as any pair of sibs. Genetic characters should therefore show a higher
concordance in MZ than DZ twins, and many characters do. Higher concordances in MZ
compared with DZ twins might also be seen if the character is determined by environmental
factors. For a start, half of DZ twins are of unlike sex, whereas all MZ twins are the same sex.
Even if the comparison is restricted to same-sex DZ twins, at least for behavioral traits the
argument can be made that MZ twins are more likely to be very similar, to be dressed and
treated the same, and thus to share more of their environment than DZ twins. MZ twins
separated at birth and brought up in entirely separate environments would provide the ideal
experiment. Such separations happened in the past more often than one might expect
because the birth of twins was sometimes the last straw for an overburdened mother.
Fascinating television programs can be made about twins reunited after 40 years of
separation, who discover they have similar jobs, wear similar clothes and like the same music.
As research material, however, separated twins have many drawbacks:
- There are very few of them, so any research is based on small numbers of arguably
exceptional people.
- The separation was often not total - often they were separated some time after birth,
and brought up by relatives.
- There is a bias of ascertainment - everybody wants to know about strikingly similar
separated twins, but separated twins who are very different are not newsworthy.
- Even in principle, research on separated twins cannot distinguish intrauterine
environmental causes from genetic causes. This may be important, for example in studies
of sexual orientation (the gay gene), where some people have suggested that maternal
hormones may affect the fetus in utero so as to influence its future sexual orientation.
Sebenta de Gentica
FCML
153
Thus, for all their anecdotal fascination, separated twins have contributed relatively little to
human genetic research.
b) Estudos de adopes
If separating twins is an impractical way of disentangling heredity from family environment,
adoption is much more promising. Two study designs are possible:
- find adopted people who suffer from a particular disease known to run in families, and ask
whether it runs in their biological family or their adoptive family;
- start with affected subjects whose children have been adopted away from the family, and
ask whether being adopted away from the family saves a child from the risk of the disease.
A celebrated (and controversial) study by Rosenthal and Kety (1968) used the first of these
designs to test for genetic factors in schizophrenia. The diagnostic criteria used in this study
have been criticized; there have also been claims (disputed) that not all diagnoses were made
truly blind. However, an independent re-analysis using DSM-III diagnostic criteria reached
substantially the same conclusions. Quadro 10 shows the results of a later extension of this
study. The main obstacle in adoption studies is lack of information about the biological family,
frequently made worse by the undesirability of approaching them with questions. A secondary
problem is selective placement, where the adoption agency, in the interests of the child,
chooses a family likely to resemble the biological family. Thus organizing a study may be
difficult. Nevertheless, in countries where efficient adoption registers exist, this is
unquestionably the most powerful method for checking how far a character is genetically
determined. Quantitative characters can be similarly investigated by comparing the correlation
between an adoptee and his/her biological or adoptive relatives.
Quadro 12: Results fot the test for genetic factors in schizophrenia
Marcadores annimos e desequilbrio de linkage
Os estudos de ligao gnica permitem identificar a presena de alelos que podem ser
herdados em conjunto com um polimorfismo de DNA que funcione como marcador, pela
proximidade entre ambos. Constituem uma forma indirecta de estudar o genoma. Para os
Sebenta de Gentica
FCML
154
estudos de ligao gnica podem ser usados os SNPs, os VNTRs e os RFLPs. Os
polimorfismos do DNA so informativos quando evidenciam heterozigotia (diferena entre as
sequncias estudadas de um par de cromossomas homlogos).
Em estudo de famlias, pode fazer-se a deteco e medio de linkage:
- pelo menos um dos progenitores tem de ser heterozigtico em ambos os loci, de modo que
a recombinao entre os loci seja reconhecvel;
- os loci mais informativos so os altamente polimrficos (como o HLA no brao curto do
cromossoma 6), onde cada progenitor possui, regra geral, 2 hapltipos diferentes;
- as famlias com mais filhos e aquelas com 3 ou mais geraes, so as mais informativas.
Uma das limitaes dos estudos de ligao gnica prende-se com a necessidade de haver
um doente na famlia j que, sem um doente, os estudos de ligao gnica no podem ser
realizados. Por isso, os resultados observados numa famlia no podem ser transferidos para
outra, uma vez que o mesmo marcador pode estar em ligao com outro alelo, com o mesmo
alelo na forma normal ou com diferentes mutaes patognicas ou no. Outras dificuldades
resultam da heterogeneidade gnica ou de uma distncia gentica grande entre o
polimorfismo marcador e o gene em ligao. Nos casos em que est presente a
heterogeneidade, a identificao da ligao gnica pode no traduzir a presena de alterao
a nvel de lcus em ligao , uma vez que o fentipo pode estar a ser determinado por outro
gene distante ou localizado noutro cromossoma.
Meiotic or linkage analysis using multiply affected and nuclear families is a potentially
powerful tool for identifying the genetic susceptibility loci of complex diseases such as asthma,
diabetes and the major psychoses. However, the results of these analyses for complex
disorders are difficult to interpret.
For schizophrenia and bipolar disorder, several loci,
including chromosomes 13q and 6p, meet the criteria for significance of linkage proposed by
Lander and Kruglyak, but the validity of many other loci remains uncertain. Power analysis
indicates that linkage may reliably detect loci conferring a relative risk of four, but may miss
loci of more modest relative risk, such as two or less. Even when the stringent criteria for
significant linkage are met, the regions implicated are typically too broad (~20 cM or more) to
attempt the analysis of candidate genes in the region. Linkage disequilibrium (LD) mapping
presents a powerful alternative to linkage analysis for the mapping and detection of genes
with a modest effect on risk. LD is a complex phenomenon and it can be affected by many
factors related to the evolutionary history of the population, including random genetic drift,
founder effects, mutation, selection, population admixture and stratification. Extension of LD
mapping to include haplotypes, which provides more information on evolutionary history of the
test population, can greatly enhance LD mapping. For complex diseases, LD mapping at
Sebenta de Gentica
FCML
155
present has largely been restricted to the use of single microsatellite markers in large
populations, but the markers of choice in future are likely to be SNPs since these are both
abundant and highly stable. Two broad approaches to the use of SNPs in LD mapping have
been suggested, either the use of a genome-wide set of anonymous markers or a catalogue
of SNPs for all coding regions. Recent sequencing analysis of the lipoprotein lipase gene,
which detected 88 variants in a 9.7-kb region, indicates that dense SNP maps within and
around genes may be required for the reliable detection of LD, rather than a genome-wide
anonymous SNP map which would mark most genes only once.
The advantage of the former
method is that the substantial additional information from multiple marker haplotypes can be
used.
LD is widely distributed in anonymous regions of the human
genome and its use may allow
more accurate measurement of small
genetic distances than does standard linkage analysis.
A GENTICA DO CANCRO
1. Mutao somtica e cancro__________________________________
Oncogenes e genes supressores de tumores
O On nc co og ge en ne es s
Os oncogenes so genes que afectam o crescimento e desenvolvimento celulares normais.
H dois tipos de oncogenes:
a) Proto-oncogenes (c-onc) - Genes semelhantes aos oncogenes virais que quando
activados provocam malignizao. Normalmente existem no genoma em diferentes locais,
sem provocar neoplasia.
b) Oncogenes virais (v-onc) - Presentes nos genomas dos vrus e uma vez incorporados no
DNA da clula infectada podem expressar-se, tornando a clula neoplsica (retrooncogenes).
A grande maioria dos oncogenes so constitudos por formas modificadas e activas de genes
normais, designados por proto-oncogenes, que esto envolvidos no controlo da proliferao e
diferenciao celular e se encontam em todas as clulas normais. Os oncogenes possuem
um efeito dominante ao nvel celular, ou seja, quando activados um nico alelo mutante
suficiente para alterar o fentipo de uma clula normal para maligna. Encontram-se
nomalmente silenciados aps terem sido muito importantes no desenvolvimento embrionrioe
a sua activao posterior provoca a sua transformao em oncogenes activos.
A activao dos proto-oncogenes, pode dever-se a vrios factores:
Sebenta de Gentica
FCML
156
1. Mutao de uma nica base - Alterao nos oncogenes ou nos factores que os regulam
por aco de agentes mutagnios externos (ex: proto-oncogene ras, que se converte a
oncogene por uma mutao pontual).
2. Translocaes cromossmicas - Translocaes de oncogenes para zonas onde deixam
de estar reprimidos, podendo ficar sob a aco de genes reguladores que os activam (ex:
enhancer, TATA box), ou libertos da influncia de sequncias reguladoras normais.
(Ex:. Um exemplo a leucemia mielide crnica, que se caracteriza pela translocao de
parte do proto-oncogene abl, da sua posio normal no cromossoma 9 para o cromossoma
22, onde fica fundido com o gene bcr, formando-se um gene quimrico com actividade
oncognica.)
3. Insero de oncogenes virais - pr-oncovrus - Ocorre na cancerignese viral. H
transcrio para o DNA celular de um segmento de DNA viral (provrus), atravs da aco da
transcriptase inversa. Como consequncia, os proto-oncogenes adjacentes a estes
segmentos de origem viral so activados (pela presena de promotores virais e alteraes
gnicas estruturais).
4. Amplificao gnica - Traduz-se na expresso excessiva de um determinado gene,
correspondendo ao que acontece em alguns tumores. Deve-se introduo de novos
segmentos ou duplicao, aquando da replicao do DNA. A gravidade do tumor tanto
maior quanto maior for o grau de amplificao. A amplificao ou reduplicao de genes,
pode ser detectada citogeneticamente pela presena de double minutes ou pela presena de
regies homogneas num cromossoma- HSR (homogenous staining regions).
5. No dissociao mittica
Os oncogenes sintetizam protenas, exercendo uma actividade desregulada sobre a
proliferao celular normal:
- Promovem a sntese de determinadas protenas intervenientes nas vias metablicas
relacionadas com o crescimento celular, que assim progride indefinidamente, sem
necessidade de estmulos.
- As protenas que codificam podem ser, por exemplo, factores de crescimento ou enzimas
chave (ex: adenilciclase) - estimulao autcrina das prprias clulas tumorais.
- Os produtos dos oncogenes podem mediar a aco de um factor de crescimento,
estimulando os receptores dos factores de crescimento (certos oncogenes codificam
receptores de factores de crescimento) e tirosinas quinases (catalisam a transferncia de
Sebenta de Gentica
FCML
157
grupos fosfato para determinadas molculas proteicas activando-as e induzindo, assim,
uma cascata de transduo de sinal no interior da clula).
- GTP binding-proteins, actuam na transduo do sinal de membrana. o caso da protena
p21, protena membranar que fixa o GTP. Quando um determinado estmulo exterior
recebido ao nvel da face externa da membrana plasmtica (havendo formao do
complexo ligando-receptor) a p21 liberta o GDP e liga-se a um GTP, que a activa. A p21
vai ento activar a adenil ciclase, com consequente aumento dos nveis intracelulares de
cAMP, que, por sua vez, ir activar as enzimas quinases dependentes do cAMP. A
actividade da p21 transitria, uma vez que ela prpria hidroliza o GTP a GDP tornando-
se novamente inactiva. A converso do proto-oncogene ras em oncogene vai alterar esta
ltima propriedade da p21, pelo que a nova protena mutada, hidrolisando o GTP com
muito menos eficcia, vai manter-se activa durante mais tempo, fornecendo uma
estimulao contnua clula.
G Ge en ne es s s su up pr re es ss so or re es s d de e t tu um mo or re es s
Os genes supressores tumorais so divididos em dois grandes grupos: os Gatekeepers e os
Caretakers. Os genes supressores de tumores carecem de homozigotia para que os seus
efeitos na desregulao da proliferao se manifeste (isto so recessivos, ao contrrio dos
oncogenes, que actuam em dominncia). A aquisio dessa homozigotia ocorre segundo o
mecanismo two hit, isto , uma mutao herdada e outra adquirida por LOH (mutao ou
deleco): o que ocorre genericamente nos cancros hereditrios
1) GATEKEEPERS OU GENES PROTETORES: regulam directamente o ciclo celular. So
genes de suscetibilidade para cancro.
- Gene p53: presente no cromossoma 17, este gene est mutado em cerca de 2/3 dos
casos de cancro. Ele responsvel pela interrupo do ciclo celular na fase G1 quando
h qualquer alterao na sequncia de DNA, a fim de que o dano seja reparado. Se o
reparo no for feito, o gene induzir a activao do mecanismo de apoptose. A disfuno
desse gene faz com que o ciclo celular prossiga mesmo que haja uma mutao no DNA,
permitindo a sua transmisso s clulas descendentes e iniciando um processo
neoplsico. Um exemplo de doena causada por alteraes no p53 a Sndrome de Li-
Fraumeni, condio em que h uma predisposio a desenvolver cancro em vrios locais,
como mama, clon, pncreas, entre outros.
- Gene RB1: situado no cromossoma 13, produz uma protena que bloqueia o ciclo celular
quando hipofosforilada. Nesta forma, a protena pRB liga-se e inactiva o factor de
transcrio E2F (que quando no ligado RB1 estimula a sntese de vrias outras
Sebenta de Gentica
FCML
158
protenas necessrias continuidade do ciclo celular). Quando o RB1 est mutado, o seu
produto encontra-se permanentemente hiperfosforilado, permitindo a progresso do ciclo
e dando incio a um processo neoplsico. Apesar desse gene se expressar em vrios
tecidos alm da retina, a sua mutao resulta geralmente num retinoblastoma, hereditrio
(em 40% dos casos) e espordico (em 60% casos).
- Gene APC: est localizado no cromossoma 5 e produz a protena apc, que regula a
quantidade de |-catenina livre no citoplasma. Em condies normais, quando a clula no
precisa se multiplicar, a |-catenina encontra-se ligada a E-caderina, inibindo a progresso
do ciclo celular. Se o gene APC estiver mutado, produzir uma protena truncada,
responsvel por um aumento da poro livre de |-catenina, que transportada para o
ncleo, activando a transcrio de genes de proliferao celular, incluindo o MYC.
Mutaes no APC provocam polipose intestinal adenomatosa de carter familiar ou
espordico e sndromes que envolvem cancro colorretal, como a sndrome de Gardner.
2) CARETAKERS OU GENES DE MANUTENO: actuam reparando danos no DNA,
mantendo a integridade genmica e evitando a instabilidade gentica. Sozinhos no induzem
a formao de neoplasias, pois as alteraes nesses genes no conferem vantagens
proliferativas clula, mas facilitam a ocorrncia de mutaes nos genes gatekeepers, as
quais daro incio cancerignese.
- Genes BRCA1 e BRCA2: esto presentes nos cromossomas 17 e 13, respectivamente.
So activados nas fases G1 e S do ciclo celular. Os produtos dos dois genes esto num
mesmo complexo multiprotico (BASC) e so responsveis pela resposta celular s quebras
do DNA que ocorrem normalmente na recombinao homloga ou de forma anormal quando
h danos na estrutura do DNA. Se mutados, predispem ao aparecimento de cancro de
mama e de ovrio, que tanto podem ter carcter espordico quanto hereditrio.
- Genes MMR: so genes responsveis por reparar erros de emparelhamento do DNA
(mismatch repair genes). H inmeros genes de reparao existentes, mas apenas alguns j
foram identificados como causadores de tumores como: MLH1, MSH2, PMSL1, PMSL2 e
MSH6. Mutaes nesses genes provocam um aumento da incidncia de mutaes pontuais
no DNA e tendncia instabilidade dos microssatlites. Essa instabilidade denominada de
fentipo Erro de Replicao Positivo (RER+), que ocorre em vrios tipos de tumores.
Alteraes nos genes de reparao provocam, mais frequentemente, cancro colorretal
hereditrio sem polipose, mas tambm so responsveis por cancros intestinais espordicos.
Sebenta de Gentica
FCML
159
Chromosome instability (CIN): Disruption of mitotic checkpoint assembly genes can lead to
CIN because checkpoint defective cells can complete mitosis with inappropriately aligned
chromosome. Recent data strongly support the hypothesis that carcinogen exposure
determines the type of instability in MIN and CIN cancers (Bardelli et al., 2001). Despite these
findings, the mechanism of CIN progression remains unclear.
O carcter multifsico e multifactorial da cancerignese. Hipteses gentica e
epigentica
C Ca ar r c ct te er r m mu ul lt ti if f s si ic co o e e m mu ul lt ti if fa ac ct to or ri ia al l d da a c ca an nc ce er ri ig g n ne es se e
O cancro um dos problemas mais comuns e severos da clnica de hoje em dia. As
estatsticas mostram que o cancro mata cerca de 1/3 da populao, responsvel por 20%
das mortes e nos pases industrializados responsvel por 10% do custo total da sade. O
cancro no uma doena nica existindo uma grande variedade de tumores malignos que
tm origem no mesmo processo de crescimento descontrolado. Um tumor um clone de
clulas provenientes de uma nica clula ancestral, na qual ocorreu o evento desencadeador
que levou hiperproliferao dessas clulas. Uma neoplasia , assim, definida como em
excessiva e desregulada proliferao celular, em competio com os tecidos normais que se
mantm, mesmo aps o estmulo que lhe deu origem ter cessado. Para alm desta
caracterstica hiperproliferao, uma neoplasia apresenta ainda invasividade e capacidade de
metastizao. O conceito de cancro como doena gentica relativamente recente. Cerca de
5% de todos os cancros parecem seguir um padro familiar na sua maioria, contudo, a
hereditariedade no parece desempenhar papel algum na maioria dos casos. No entanto
muitos cancros possuem uma componente gentica significativa pois certas pessoas so
mais susceptveis a determinada neoplasia maligna, em consequncia de defeitos genticos
que predispem ao cancro. Apesar da natureza multifactorial do cancro (ambiencial, gentica,
imunolgica, etc) h um denominador comum a todas as neoplasias: a alterao do DNA.
Toma-se, assim, evidente que todos os cancros resultam de mutaes em clulas somticas
e que a sua progresso envolve a expresso de uma srie de genes. As mutaes que levam
ao cancro afectam os genes relacionados com a proliferao e desenvolvimento celulares. A
proliferao celular clonal, isto , provm de uma nica clula transformada, em que todas
as clulas filhas so iguais progenitora. No entanto, o crescimento e multiplicao destas
clulas to rpido que os mecanismos de sntese do DNA, por vezes, no conseguem
acompanhar e surgem outras anomalias cromossmicas (deleces, translocaes,
duplicaes). Surgem ento novos mutantes que podem ter dois destinos: so inviveis e
morrem ou formam novos subclones com propriedades diferentes. O primeiro passo para a
carcinognese a leso do DNA, quer devido a factores ambienciais, quer devido a factores
Sebenta de Gentica
FCML
160
endgenos. Esta leso pode levar morte celular, pode ser reparada (caso os mecanismos
de reparao estejam ntegros) ou pode levar fixao de mutaes. As mutaes podem
ser deletrias (levando morte celular), vantajosas (promovendo vantagem adaptativa) ou
silenciosas. Se a mutao lesar um anti-oncogene, um oncogene ou um gene regulador da
apoptose, h crescimento rpido e descontrolado das clulas anormais (cancergenas), as
quais tm e adquirem alteraes cromossmicas, morfolgicas e funcionais. Ocorre assim
proliferao de clulas neoplsicas e crescimento tumoral. As mutaes podem ocorrer em
qualquer clula, tanto na linhagem germinativa como na somtica. As mutaes somticas,
das quais resulta um mosaicismo somtico, podem originar uma proporo significativa de
cancros. A mutao que leva perda ou alterao da funo de um gene pode ser
considerada do ponto de vista da adaptabilidade, que determina se uma mutao se perde
imediatamente, se sobrevive algumas geraes ou se torna um alelo predominante no locus
envolvido. A adaptabilidade de um alelo representa um equilbrio entre a taxa de mutao
naquele gene e os efeitos de seleco (sendo a taxa de seleco igual mutao de um
gene /diviso celular). A incidncia de neoplasias aumenta com a idade. Os agentes
indutores de neoplasias podem ser divididos em genotxicos, mutagnios e clastognicos.
Os genotxicos alteram o DNA e protenas afins, os mutagnios causam alterao da
estrutura primria do DNA, com fixao de uma mutao, os clastognicos provocam
alteraes cromossmicas estruturais, induzindo a ruptura de pores de cromossomas
atravs da quebra de ligaes (ex: radiaes ionizantes).
Two distinct pathways of tumorigenesis exist in sporadic colorectal cancer:
Chromosome instability (CIN): Disruption of mitotic checkpoint assembly genes can lead to
CIN because checkpoint defective cells can complete mitosis with inappropriately aligned
chromosome. Recent data strongly support the hypothesis that carcinogen exposure
determines the type of instability in CIN cancers. Despite these findings, the mechanism of
CIN progression remains unclear.
Microsatellite instability (MIN): is characterized by widespread microsatellite instability due
to aberrant mismatch repair machinery
H Hi ip p t te es se es s g ge en n t ti ic ca as s
Os tumores resultam de anomalias genticas congnitas ou induzidas, que ditam o
comportamento aberrante das clulas. Cerca de 5% dos cancros parecem seguir um padro
familiar; aparentemente nos restantes a herana tem pouca ou nenhuma participao.
Contudo, muitos cancros apresentam caractersticas de herana multifactorial, possuindo um
componente gentico significativo (predisposio para determinada neoplasia). Tornou-se
Sebenta de Gentica
FCML
161
evidente que todos os cancros, mesmo na ausncia de qualquer componente hereditrio
aparente, resultam de mutaes somticas e que a sua progresso tambm envolve a
expresso de uma srie de genes.
H Hi ip p t te es se e e ep pi ig ge en n t ti ic ca a
O comportamento das clulas tumorais resulta da expresso de genes no mutados, que
esto desregulados ou anormalmente controlados. As principais classes de agentes
cancergenos incluem:
- radiaes (U.V., X e ); compostos qumicos provenientes da dieta, profisso (ex:
pesticidas), vcios (tabaco - hidrocarbonetos aromticos policiclcos), medicao; agentes
biolgicos; oncovrus (ex: hepatite B - cancro heptico; vrus de Epstein-Barr - linfoma de
Burkit, HTLV-1 - leucemia das clulas T; herpes simplex - cancro do tero);
Os genes que predispem para a ocorrncia de cancro podem actuar de vrios modos:
1. Afectar a velocidade a que substncias pr-carcinognicas exgenas so metabolizadas
em carcinognios activos, que podem danificar directamente o genoma.
2. Afectar a estabilidade cromossmica e a capacidade do organismo para reparar o DNA
lesado.
3. Afectar a vigilncia do sistema imunitrio, diminuindo a capacidade do organismo para
reconhecer e eliminar clulas tumorais.
4. Afectar os mecanismos de regulao da proliferao e diferenciao celular.
O desenvolvimento de uma neoplasia divide-se nas seguintes fases:
I Iniciao
Fase que ocorre rapidamente, estando envolvida uma leso irreversvel do DNA num gene
crtico que codifique uma protena integrada numa via que leva proliferao celular ou ao
bloqueio desta. Isto resulta da aco de um agente carcinognico que actua no genoma
celular. O agente que despoleta a cancerignese denominado de iniciador. No conduz ao
aparecimento de tumor se no activar a promoo.
II Promoo
Ocorre por aco dos agentes promotores cuja aco amplificar e acelerar o efeito do
iniciador. Estes compostos ligam-se no covalentemente, de modo reversvel, a um receptor
de membrana, estimulando vias intracelulares de transduo de sinal, que mimetiza um sinal
mitognico deixando de haver possibilidade de controlo sistmico da proliferao local. Os
agentes promotores no so mutagnicos e a exposio das clulas a estes agentes tem de
ser prolongada, da que este estadio da cancerignese seja geralmente lento (meses, anos).
Sebenta de Gentica
FCML
162
III Progresso ou proliferao
Desenvolvimento da massa tumoral. Novos clones podem surgir por novas mutaes no
clone primrio. Estes podem possuir caractersticas que lhes proporcionem melhor adaptao
ao meio e vantagens face a certas caractersticas desse meio. Neste estadio as clulas
tumorais apresentam um crescimento autnomo.
IV - Metastizao ou generalizao-
"Colonizao distncia" do tumor e invaso dos tecidos adjacentes.
Fig. 70: Etapeas da carcinognese
Origem monoclonal vs. policlonal dos tumores
Cancer cells are indigenous cellsabnormal cells that arise from the body's normal tissues.
Furthermore, virtually all malignant tumors are monoclonal in origin, that is, derived from a
single ancestral cell that somehow underwent conversion from a normal to a cancerous state.
How do we know that tumors are monoclonal? Two distinct scenarios might explain how
cancers develop within normal tissues. In the first, many individual cells become cancerous,
and the resulting tumor represents the descendants of these original cells. In this case, the
tumor is polyclonal in nature. In the second scenario, only one cell experiences the original
transformation from a normal cell to a cancerous cell, and all of the cells in the tumor are
descendants of that cell. Direct evidence supporting the monoclonal origin of virtually all
Sebenta de Gentica
FCML
163
malignant tumors has been difficult to acquire because most tumor cells lack obvious
distinguishing marks that scientists can use to demonstrate their clonal relationship. There is,
however, one cellular marker that scientists can use as an indication of such relationships: the
inactivated X chromosome that occurs in almost all of the body cells of a human female. X-
chromosome inactivation occurs randomly in all cells during female embryonic development.
Because the inactivation is random, the female is like a mosaic in terms of the X chromosome,
with different copies of the X turned on or off in different cells of the body. Once inactivation
occurs in a cell, all of the future generations of cells coming from that cell have the same
chromosome inactivated in them as well (either the maternal or the paternal X). The
observation that all the cells within a given tumor invariably have the same X chromosome
inactivated suggests that all cells in the tumor must have descended from a single ancestral
cell.
Oncogene addiction. Passenger and Driver mutations
Oncogene addiction:
Traduz o facto de que embora possa haver mutaes em mltiplos genes com papel na
cancerignese (driver mutations), um s gene pode condicionar por aco em rede a diviso
celular e a proliferao, o que tem evidentes consequncias teraputicas.
Cancer cells contain multiple genetic and epigenetic abnormalities.
Despite this complexity,
their growth and survival can often
be impaired by the inactivation of a single oncogene. This
phenomenon,
called "oncogene addiction," provides a rationale for molecular
targeted therapy.
The efficacy of this strategy requires novel
methods, including integrative genomics and
systems biology,
to identify the state of oncogene addiction in specific cancers. Combination
therapy may also be
required to prevent the escape of cancers from a given state
of oncogene
addiction.
Sebenta de Gentica
FCML
164
We have previously proposed that the phenomenon of oncogene addiction is a
consequence of the fact that the multistage process of carcinogenesis is not simply a
summation of the individual effects of activation of multiple oncogenes and inactivation of
multiple tumor suppressor genes. This proposal is consistent with the fact that the proteins
encoded by these genes often have multiple roles in complex and interacting networks, which
display both positive and negative feedback control. The function of these proteins is also
influenced by their levels of activity and the context in which they are expressed. Thus, a
given oncogene can enhance cell proliferation but it can also enhance apoptosis.
Furthermore, throughout the multistage carcinogenic process, the evolving cancer cell must
maintain a state of homeostasis between positive-acting and negative-acting factors in order
to maintain structural integrity, viability, and the capacity to replicate. For these reasons, the
intracellular circuitry or 'wiring diagram' that regulates signal transduction and gene expression
in cancer cells is very different, i.e., 'bizarre,' when compared to that of normal cells.
In cancer
cells a given oncogene may play a more essential and qualitatively different role in a given
pathway or 'module' compared with its role in normal cells. Thus, cancer cells may be much
more dependent on the activity of a specific oncogene than normal cells. Within the context of
disordered cell circuitry, specific mechanisms have been proposed to explain why inactivation
of an oncogene might lead to selective growth inhibition, differentiation and/or apoptosis in
cancer cells but not in normal cells that express the same oncogene. One explanation is that,
in order to maintain homeostasis, the proliferation-enhancing effects of a specific oncogene in
cancer cells might be partially buffered through negative feedback mechanisms, through
increased expression of proliferation-inhibitory factors.
If this oncogene is then inactivated the
cancer cells might suffer a relative excess of the latter inhibitory factors and thus undergo
apoptosis, before a new level of homeostasis can be achieved. The apparent propensity of
some cancer cells to undergo apoptosis when stressed
could enhance this process. A second
mechanism is based on the concept of 'synthetic lethality' originally derived from studies in
lower organisms. According to this concept, two genes are said to be synthetic lethal if
mutation of one of the two genes is compatible with survival but mutation of both genes
causes cell death.
For example, certain cancer cells might be highly dependent on a given
oncogene because during their development they lost the function of another gene that
normally performs a similar function. A drug that inhibits the activity of the oncogene would,
therefore, selectively target the cancer cells and spare the normal cells. Furthermore, because
of the bizarre circuitry of cancer cells, pairs of genes in cancer cells that have a synthetic
lethal relationship may differ from those in normal cells, thus increasing the dependence of
tumor cells on a specific oncogene. A related explanation for oncogene addiction is that,
during the multistage carcinogenesis process, cancer cells become highly dependent on
Sebenta de Gentica
FCML
165
specific oncogenes and their related pathways because of the large numbers of mutated and
inactivated genes that normally function in other pathways. This dependence could render
cancer cells less adaptable than normal cells.
Passenger and Driver mutations
All cancers arise as a result of somatically acquired changes in the DNA of cancer cells. That
does not mean, however, that all the somatic abnormalities present in a cancer genome have
been involved in development of the cancer. Indeed, it is likely that some have made no
contribution at all. To embody this concept, the terms 'driver' and 'passenger' mutation have
been coined. A driver mutation is causally implicated in oncogenesis. It has conferred growth
advantage on the cancer cell and has been positively selected in the microenvironment of the
tissue in which the cancer arises. A driver mutation need not be required for maintenance of
the final cancer (although it often is) but it must have been selected at some point along the
lineage of cancer development shown in fig. 70. A passenger mutation has not been selected,
has not conferred clonal growth advantage and has therefore not contributed to cancer
development. Passenger mutations are found within cancer genomes because somatic
mutations without functional consequences often occur during cell division. Thus, a cell that
acquires a driver mutation will already have biologically inert somatic mutations within its
genome. These will be carried along in the clonal expansion that follows and therefore will be
present in all cells of the final cancer. Some somatic mutations may actually impair cell
survival. These will usually be subject to negative selection and hence be absent from the
cancer genome.
Fig. 71: Mutations may be acquired while the cell lineage is phenotypically normal, reflecting both the intrinsic
mutations acquired during normal cell division and the effects of exogenous mutagens. During the development of
the cancer other processes, for example DNA repair defects, may contribute to the mutational burden. Passenger
mutations do not have any effect on the cancer cell, but driver mutations will cause a clonal expansion. Relapse
after chemotherapy can be associated with resistance mutations that often predate the initiation of treatment.
Sebenta de Gentica
FCML
166
Leucemia mielide crnica e tirosina-quinases. Receptores de transporte e
resistncia teraputica
A leucemia mieloide crnica (LMC) uma neoplasia da medula ssea que transforma a
clula progenitora hematopotica normal em maligna.
ocasionada por uma translocao
entre os cromossomos 9 e 22 t(9:22)(q34;11),
determinando assim a presena do
cromossoma Philadelfia (Ph
+
), que resultante da juno da regio do gene BCR do
cromossoma 22 com o gene ABL do cromossoma 9, gerando o gene hbrido BCR-ABL no
cromossoma 22 q-. O gene hbrido BCR-ABL produz uma protena quimrica com actividade
tirosina quinase elevada que responsvel pela patognese da LMC. O resultado desta
actividade elevada induz a medula ssea a proliferar um clone de clulas mieloides malignas
constantemente, resultando num nmero excessivo destas clulas.
Em condies normais, o
gene BCR expresso no cromossoma 22 codifica uma protena com funo relacionada
regulao do ciclo celular, enquanto o gene ABL expresso no cromossoma 9 codifica uma
protena tirosina quinase.
O gene recproco, resultante da translocao ABL-BCR no
cromossomo 9q
+
, apesar de activo, no possui nenhum papel na etiologia de doenas. Por
ser uma enzima que participa no metabolismo celular, a tirosina quinase est relacionada
com vrias patologias.
Fig. 72: cromossoma Ph
+
Os mecanismos principais resultantes da mieloproliferao constante das clulas malignas
so a alterao da adeso das clulas progenitoras s clulas estromais e matriz
extracelular, a manuteno de um sinal mitognico constante e a resistncia apopttica
celular. A enzima tirosina quinase Abl confere clula leucmica alta resistncia morte
celular independentemente do agente indutor deste processo.
Os mecanismos de resistncia
que favorecem o crescimento e a vantagem proliferativa celular Ph
+
em relao s clulas
normais no esto totalmente elucidados. Quando as clulas malignas so geradas e a
funo normal no organismo no exercida, temos como resultado os tumores que so
Sebenta de Gentica
FCML
167
ocasionados pela proliferao, adeso, apoptose e diferenciao inadequadas das clulas
malignas. Inibidores das enzimas tirosina quinases levam a clula maligna a entrar em
apoptose, reduzindo assim a proliferao tumoral.
Estudos tm evidenciado o papel essencial
do controle da actividade da tirosina quinase Abl para o tratamento da LMC.
Novas drogas
esto a ser desenvolvidas para que sejam utilizadas em doentes nos quais o tratamento
convencional tenha falhado ou que os efeitos adversos intensos tenham sido observados. A
composio destas novas drogas baseia-se na inibio especfica da expresso gnica da
doena e tem a funo de bloquear a actividade quinase da protena Bcr/Abl levando
remisso da LMC. A LMC subdividida de acordo com os exames clnicos e laboratoriais em
fase inicial crnica, fase acelerada duradoura e fase final aguda.
Nesta ltima fase pode
ocorrer a crise blstica, que considerada a mais grave.
No incio, a LMC de progresso
lenta; sendo assim, a especificidade do tratamento para estes pacientes importante, uma
vez que esta doena pode evoluir para as fases acelerada e aguda, colocando em risco a
vida do doente. Os passos iniciais da terapia para LMC so a estabilizao das clulas
sanguneas, a resposta hematolgica e citogentica. A resposta hematolgica inclui reduo
na contagem absoluta de clulas brancas, eliminao de clulas mieloides imaturas do
sangue perifrico e erradicao dos sinais e sintomas da doena. A resposta citogentica
baseada na reduo ou eliminao de clulas do cromossomo Ph
+
.
As terapias convencionais
para o controle da LMC so a quimioterapia com hidroxiureia, terapias medicamentosas com
interferron-alfa (INF-), bussulfano ou citarabina em baixa dose, a infuso linfocitria e o
transplante de clulas medulares. O tratamento da LMC avanou consideravelmente nos
ltimos anos e a terapia molecular com inibidores de tirosina quinase mudou drasticamente a
terapia convencional, demonstrando resultados promissores para os doentes.
2. Sndromes de cancro familiar________________________________
Mutaes nos genes BRCA1 e BRCA2 no cancro da mama familiar
Cancro da mama familiar (forma Mendeliana do cancro da mama):
- surge mais precocemente que o espontneo
- responsvel por cerca de 10% dos cancros da mama
- dois genes envolvidos: Brca1 (17q21) Brac2 (13q12) heterogeneidade de lcus
- cada um destes genes apresenta tambm heterogeneidade allica.
Quanto ao Brca1 e Brca2:
- ambos so classificados como genes supressores de tumores envolvidos:
Sebenta de Gentica
FCML
168
1. na reparao do DNA. No ncleo de muitas clulas as proteinas do BRCA1 e BRCA2
interagem com o RAD51 (e seus parlogos XRCC2 e XRCC3) para reparar o DNA quando h
quebras da dupla cadeia de DNA ajudando a manter a sua estabilidade.
Fig. 73: BRAC1 e BRAC2 na reparao do DNA
O BRCA1 tambm promove reparaes do DNA atravs de NHEJ (non homologous end
joining) por inibir a nuclease do complexo MRN. Na ATM (ataxia telangiectasia) a cinase
activada por radiao ionizante fosforila BRCA1 e FANCD2. Na NBS (Nijmegen Breakage
Syndrome) - NBS1 tem duas funes: induz paragem em S e interage com BRCA1 na
reparao de DNA. Na FANC (anemia de Fanconi) - FancA, FancC, FancE, FancF, e FancG
formam um complexo que interage com DNA e levam a monoubiquitinao de FANCD2.
Atravs de associao com BRCA1 e BRCA2 leva a reparao de DNA.
Fig. 74: Interaco do BRAC1 e 2 com Fanc, NBS e ATM
Sebenta de Gentica
FCML
169
2. Ubiquitinizao
3. Regulao transcriptional
4. Controlo do ciclo celular. O BRCA 1 e 2 relacionam-se com o BASC = BRCA-associated
genome surveillance complex na regulao do cilco celular.
- quando h mutaes h probabilidade de desenvolver tumores (mama e ovrio
essencialemente)
Fig. 75: Mutaes no gene BRAC1/BRAC2
- mutao no Brca1: probabilidade >50% (acima dos 60anos) de desenvolver cancro da
mama e 20% de probabilidade de desenvolver cancro do ovrio
-mutao no Brca2: um bocado menos que brca1 na mama mas no ovrio muito reduzido
Fig. 76: BRCA 1 Fig. 77: BRCA2
(^ risco de cancro de mama; risco de cancro de ovrio)
Mutao no gene RB e retinoblastoma hereditrio por perda de heterozigotia
um tumor embrionrio da retina em que se observa uma deleco em 13q14. A penetrncia
do gene do retinoblastoma hereditrio (uni ou bilateral) de cerca de 80%. Quanto aos casos
isolados de retinoblastoma unilateral, estimou-se que apenas 20% devem ser hereditrios. O
gene RB1, produz uma protena que bloqueia o ciclo celular quando hipofosforilada. Nesta
Sebenta de Gentica
FCML
170
forma, a protena pRB liga-se e inactiva o factor de transcrio E2F (que quando no ligado
RB1 estimula a sntese de vrias outras protenas necessrias continuidade do ciclo
celular). Quando o RB1 est mutado, o seu produto encontra-se permanentemente
hiperfosforilado, permitindo a progresso do ciclo e dando incio a um processo neoplsico.
Fig. 78: Rb e a expresso gentica
A hiptese two hit para explicar a manifestao de retinoblastomas, assume que ambos os
alelos normais do locus Rb tm de ser inactivados para que se desenvolva o tumor (como
acontece com outros genes supressores de tumores). O gene encontra-se localizado no
cromossoma 13 (13q14). Na forma familiar, a criana nasce com uma cpia normal e outra
alterada do gene Rb first hit. Atravs de uma mutao somtica (mutao pontual, deleco
intersticial, ou mesmo completa, do cromossoma 13) perde-se a cpia intacta (normal) desse
gene second hit. The most frequent second hit in retinoblastoma patients is loss of
heterozygosity (or LOH: represents the loss of normal function of one allele of a gene in which
the other allele was already inactivated) followed by promoter methylation. Esta 2 mutao
pode acontecer numa ou em vrias clulas da retina, podendo portanto, desenvolver-se
vrios focos tumorais em ambas as retinas.
Fig. 79: Hiptese de two-hit de Knudson para explicar o aparecimento de retinoblastia espordica. Suponhamos
que temos um milho (10
6
) de clulas alvo e que a probabilidade de mutao de 10
-5
por clula. No
retinoblastoma espordico necessrio 2hits para afectar 1 pessoa em 10 000 enquanto que no retinoblastoma
hereditrio o 1 hit herdado logo s necessrio o segundo para que doena tenha elevada penetrncia.
Sebenta de Gentica
FCML
171
Polipose adenomatosa familiar (FAP) e o sndrome de Lynch (HNPCC)
A FAP e o HNPCC so responsveis por 10% dos cancros do clon.
A FAP uma doena autossmica dominante na qual o clon forrado com centenas de
milhares de plipos. Estes plipos (adenomas) no so malignos mas se no forem
removidos quase certo que um dia evoluiro para carcinoma invasivo. Esta doena deve-se
a uma mutao do gene APC situado no cromossoma 5q21. Mais de 90% das mutaes do
gene APC, nos tumores colorectais, resulta numa protena APC truncada, devido a mutaes
frameshift ou nonsense. Tambm podem ocorrer mutaes que alteram o splicing. Na
FAP, a perda contitucional de um exemplar do gene APC no 5q21 suficiente para revestir o
colon com plipos adenomatosos. No h expresso do APC nos FAP detectveis mesmo
numa fase muito inicial. Cerca de uma clula epithelial em 10
6
desenvolve um plipo uma
taxa consistente com a perda do segundo alelo APC (pode ser tambm devido perda de
heterozigotia) determinante no processo de desenvolver cancro.
Fig. 80: Processo da FAP
A protena APC contm diversos domnios funcionais, que incluem locais de ligao aos
microtbulos e de ligao e degradao da -catenina, que mantm estacionrios os nveis
de -catenina citoplasmtica (estimula a transcrio)
A regio COOH- terminal de APC liga-se aos microtbulos (MTs) e promove a polimeri-
zao de tubulina como as microtubule-associated proteins (MAPs).
O produto da expresso gentica do gene APC liga-se -catenina que:
- se liga molcula de adeso celular E-caderina activando-a. A E-caderina uma protena
de adeso Ca2+-dependente, que controla a adeso entre clulas epiteliais e a motilidade no
desenvolvimento embrionrio ao afectar a migrao celular e a morfognese.
- transmite os sinais Wnt ao ncleo e interage com os factores de transcrio LEF-1/TCF-4
permitindo a transcrio de diversos genes, designadamente oncogenes. Os WNTs
Sebenta de Gentica
FCML
172
constituem uma famlia de glicoprotenas exportadas que so ligandos (parcrinos) dos
receptores transmembranrios frizzled. A cascata Wnt bloqueia intra-celularmente a
degradao da -catenina, conduzindo acumulao de -catenina. Na ausncia de sinal
Wnt, a -catenina fosforilada pela GSK3 e degradada no proteassoma, aps ubiquitinao.
Groucho repressor da transcrio mediada por TCF e -catenina.
Os dados sugerem ainda que a APC est envolvida na adeso celular. Uma possibilidade
que o complexo APC regule a transmisso do sinal de inibio por contacto na clula.
Na FAP, a ausncia de APC faz com que os nveis de -catenina aumentem, aumentando a
transcrio que leva a uma proliferao celular (que leva ao aparecimento do cancro)
Fig. 81: APC, Wnt e -catenina
Nos cancros colorectais espordicos (no hereditrios) por vezes identificvel o gene APC
intacto no entanto, so visiveis mutaes na -catenina que nexte contecto actua como um
oncogne.
Sebenta de Gentica
FCML
173
Diagnstico molecular e diagnstico pr-sintomtico (sensibilidade): anlise de linkage
(requer, pelo menos, dois membros afectados da famlia (99%)); Protein Truncation Test ou
PTT (8090%) e Sequenciao (>90%).
O HNPCC (Hereditary nonpolyposis colorectal cancer) tambm uma doena autossmica
dominante com alta penetrncia mas, contrariamente FAP no precedido de fase poliposa.
Tem uma penetrncia de 80-85%. HNPCC causado pela mutao de uma famlia de genes-
Mismatch repair (MMR)- sendo este o first hit, seguido pela inactivao pelo alelo sem
mutao (por LOH por exemplo)- second hit. Quando ambos os alelos dos genes MMR esto
inactivados (causado por mutao missense e frameshift essencialmente) o sistema de
reparao mismatch (MMR) do DNA- importante na manuteno da estabilidade genmica
durante a replicao do DNA- perde a sua capacidade. Como consequncia h uma
acumulao de mutaes no genoma tornando-o instvel em certas regies (micro-satlites
essencialmente). As mutaes no genes MMR: MLH1, MSH2 e MSH6 contribuem para 42-
75% dos casos de HNPCC.
Fig. 83 : MMR genes effects
Loss of heterozygosity studies on colon cancer produced novel and unexpected results in
some patients. Rather than lacking alleles present in the constitutional DNA, some tumor
specimens appeared to contain extra, novel, alleles of the microsatellite markers used. LoH is
a property of particular chromosomal regions, but the microsatellite instability (MSI or MIN)
seemed to be general. Tumors could be classified into MSI
+
or MSI
-
. MSI
+
tumors gained
Sebenta de Gentica
FCML
174
alleles for a good proportion of the markers tested, regardless of their chromosomal location.
Microsatellite instability is a characteristic feature of the autosomal dominant hereditary non-
polyposis colon cancer. Patients with HNPCC are constitutionally heterozygous for a loss-of-
function mutation. Their normal cells still have a functioning mismatch repair system and do
not show the MSI
+
phenotype. In a tumor, the second copy is lost. Interestingly, these tumors
are unusual in having relatively normal karyotypes - showing that while genetic instability is
important in cancer, it may be achieved either through instability at the nucleotide level or by
chromosomal instability.
P53 e sndrome de Li-Fraumeni
A protena p53 codificada pelo gene supressor de tumores TP53 localizado no cromossoma
17p12. considerada a guardi do genoma uma vez que impede a replicao do DNA
danificado detendo as clulas entre G1 e S at que o dano seja reparado. Caso o dano seja
irreparvel a p53 induz a apoptose celular evitando assim uma possvel proliferao tumoral.
Cerca de 80% das mutaes no p53 so missense. Mutaes no gene TP53, com ou sem
deleco allica, representam uma das alteraes genticas mais comuns identificadas em
tumores malignos. A grande maioria destas mutaes est entre os exes 5 e 8, em quatro
regies conservadas do gene TP53. Mais de 905 das mutaes esto localizadas entre os
codes 120 e 290- regio hotspot. Mutaes contitucionais deste gene so encontradas em
famlias com o sndrome de Li-Fraumeni (autossmico dominante). Os membros da famlia
afectados sofrem de tumores primrios mltiplos que compreendem, tipicamente, sarcomas
de tecidos moles, osteossarcomas, leucemia, tumores da mama, crebro e crtex da supra-
renal. O processo para o desenvolvimento desta sndorme envolve o modelo de two-hits de
Knudson (figura seguinte)
Fig. 84: LOH in Knudsons double-hit tumorigenesis model in Li-Fraumeni syndrome
Sebenta de Gentica
FCML
175
Haploinsuficincia e dominncia negativa (p53)
Haploinsuficiencia: Hoje existem dois modelos para explicar a carcinognese envolvendo
genes supressores tumorais. O primeiro e mais difundido a hiptese de Knudson, em que
mutaes devem provocar perda de funo dos dois alelos (mencionada anteriormente).
Essas mutaes tm carter recessivo, uma vez que um nico alelo mutado no capaz de
induzir uma neoplasia. O segundo o Modelo da Haploinsuficincia, em que apenas um alelo
mutado, associado a eventos adicionais promotores de tumor, seria capaz de induzir a
carcinognese, mesmo com o outro alelo a expressar-se normalmente. O Modelo da
Haploinsuficincia est baseado em estudos que demonstraram a ocorrncia de cncro por
alterao de genes supressores tumorais, mesmo estando presente apenas um alelo mutado.
A manuteno de um alelo selvagem no suporta o efeito do alelo mutado, quer seja por
produo de protena em nveis insuficientes, quer seja por um efeito dominante negativo, em
que o alelo mutado bloqueia a actividade da protena normal. O defeito na expresso de um
alelo contribui para a formao de tumor por conferir vantagem proliferativa clula ou por
causar instabilidade gentica. A haploinsuficincia no resulta diretamente num determinado
fentipo neoplsico, necessitando de outros eventos promotores de tumor. So relativamente
poucos os genes que apresentam haploinsuficincia.
Dominncia negativa: A dominant negative effect occurs when a mutant polypeptide not only
loses its own function, but also interferes with the product of the normal allele in a
heterozygote. Dominant negative mutations cause more severe effects than deletion or
nonsense mutations in the same gene. Some sort of physical association of the normal and
mutant products is required for a dominant negative effect. Structural proteins that contribute
to multimeric structures are vulnerable to dominant negative effects. Collagens is an example.
A presena, em heterozigotia, de uma mutao patognica do gene TP53 responsvel
pelo efeito de dominncia negativa.
Sebenta de Gentica
FCML
176
Fig. 86: Dominant negative effects of collagen gene mutations- Collagen fibrils are built of arrays of triple-helical
procollagen units. The type I procollagen comprises two chains encoded by the COL1A1 gene and one encoded by
COL1A2. Null mutations in either gene have a less severe effect than mutations encoding polypeptides which
cause the triple helix to be non-functional
MEN1 e MEN2 (multiple endocrine neoplasia type 1 and type 2)
Multiple endocrine neoplasia is part of a group of disorders that affect the body's network of
hormone-producing glands (the endocrine system). Hormones are chemical messengers that
travel through the bloodstream and regulate the function of cells and tissues throughout the
body. Multiple endocrine neoplasia involves tumors in at least two endocrine glands; tumors
can also develop in other organs and tissues. These growths can be noncancerous (benign)
or cancerous (malignant). If the tumors become cancerous, some cases can be life-
threatening. The two major forms of multiple endocrine neoplasia are called type 1 and type 2;
they are often confused because of their similar names. Type 1 and type 2 are distinguished
by the genes involved, the types of hormones made, and the characteristic signs and
Sebenta de Gentica
FCML
177
symptoms. Many different types of tumors are associated with multiple endocrine neoplasia.
Type 1 frequently involves tumors of the parathyroid gland, pituitary gland, and pancreas.
Tumors in these glands lead to the overproduction of hormones. The most common sign of
multiple endocrine neoplasia type 1 is overactivity of the parathyroid gland
(hyperparathyroidism). Hyperparathyroidism disrupts the normal balance of calcium in the
blood, which can lead to kidney stones, thinning of bones, nausea and vomiting, high blood
pressure (hypertension), weakness, and fatigue. The most common sign of multiple endocrine
neoplasia type 2 is a form of thyroid cancer called medullary thyroid carcinoma. Some people
with this disorder also develop a pheochromocytoma, which is a tumor of the adrenal glands
that can cause dangerously high blood pressure. Multiple endocrine neoplasia type 2 is
divided into three subtypes: type 2A, type 2B, and familial medullary thyroid carcinoma
(FMTC). These subtypes differ in their characteristic signs and symptoms and risk of specific
tumors; for example, hyperparathyroidism occurs only in type 2A. The features of this disorder
are relatively consistent within any one family. Each of the two major types of multiple
endocrine neoplasia affects an estimated 1 in 30,000 people. Among the subtypes of multiple
endocrine neoplasia type 2, type 2A is the most common form, followed by familial medullary
thyroid cancer. Type 2B is relatively uncommon, accounting for about 5 percent of all cases of
multiple endocrine neoplasia type 2. Mutations in the MEN1 and RET genes cause multiple
endocrine neoplasia. Mutations in the MEN1 gene cause multiple endocrine neoplasia type 1.
This gene provides instructions for producing a protein called menin. Menin acts as a tumor
suppressor, which means it normally keeps cells from growing and dividing too rapidly or in an
uncontrolled way. Although the exact function of menin is unknown, it is likely involved in cell
functions such as copying and repairing DNA and regulating the activity of other genes. If
mutations inactivate both copies of the MEN1 gene, it is no longer able to control cell growth
and division. The loss of functional menin allows cells to divide too frequently, leading to the
formation of tumors characteristic of multiple endocrine neoplasia type 1. Multiple endocrine
neoplasia type 2 results from mutations in the RET gene. This gene provides instructions for
producing a protein that is involved in signaling within cells. The RET protein triggers chemical
reactions that instruct cells to respond to their environment, for example by dividing or
maturing. Mutations in the RET gene overactivate the protein's signaling function, which can
trigger cell growth and division in the absence of signals from outside the cell. This unchecked
cell division can lead to the formation of tumors in endocrine glands and other tissues. Most
cases of multiple endocrine neoplasia type 1 are considered to have an autosomal dominant
pattern of inheritance. People with this condition are born with one mutated copy of the MEN1
gene in each cell. In most cases, the altered gene is inherited from an affected parent. The
remaining cases are a result of new mutations in the MEN1 gene, and occur in people with no
Sebenta de Gentica
FCML
178
history of the disorder in their family. Unlike most other autosomal dominant conditions, in
which one altered copy of a gene in each cell is sufficient to cause the disorder, two copies of
the MEN1 gene must be altered to trigger tumor formation in multiple endocrine neoplasia
type 1. A mutation in the second copy of the MEN1 gene occurs in a small number of cells
during a person's lifetime. Almost everyone who is born with one MEN1 mutation acquires a
second mutation in certain cells and develops tumors of the endocrine glands and other
tissues. Multiple endocrine neoplasia type 2 is also usually inherited in an autosomal dominant
pattern. A person with multiple endocrine neoplasia type 2 often inherits the altered RET gene
from one parent with the condition. Some cases, however, result from new mutations in the
gene and occur in people without other affected family members.
Alteraes cromossmicas e cancro
Deleco no cromossoma 11 e tumor de Wilms
uma neoplasia embrionria, transmitida de modo autossmico recessivo que se caracteriza
por um tumor maligno do rim (nefroblastoma), associado ou no inexistncia de ris
(aniridia). Muitos doentes apresentam outras alteraes, como atraso mental e do
crescimento e rgos genitais anormais. Da outra designao desta doena sndrome de
WAGR (Wilms Aniridia Genitalia Retardation). Evidencia-se uma microdeleco do brao
curto do cromossoma 11 d (11p13). O oncogene activado, aps deleco do seu supressor,
o c-Ha-ras. Pode ser explicado pela hiptese two hit, em que a deleco de 11p13 um
dos passos necessrios.para a manifestao do tumor, ou seja, a perda da heterozigotia
resulta numa homozigotia para o locus WAGR.
Wilms tumor, aniridia, genitourinary anomalies, and mental retardation syndrome, more
commonly known by the acronym WAGR syndrome, is a condition that affects the
development of many body systems. Most people with WAGR syndrome have aniridia, an
absence of the colored part of the eye (the iris). This can cause reduction in the sharpness of
vision (visual acuity) and increased sensitivity to light (photophobia). Aniridia is typically the
first noticeable sign of WAGR syndrome. Other eye problems may also be present, such as
clouding of the lens of the eyes (cataracts), increased pressure in the eyes (glaucoma), and
involuntary eye movements (nystagmus). Another common feature is intellectual disability
(mental retardation). Some individuals with WAGR syndrome also have psychiatric or
behavioral problems including depression, anxiety, attention deficit hyperactivity disorder
(ADHD), obsessive-compulsive disorder (OCD), or a developmental disorder called autism
that affects communication and social interaction. Abnormalities of the genitalia and urinary
tract (genitourinary anomalies) are seen more frequently in affected males than in affected
Sebenta de Gentica
FCML
179
females. The most common genitourinary anomaly in affected males is undescended testes
(cryptorchidism). Females may not have functional ovaries and instead have undeveloped
clumps of tissue called streak gonads. Females may also have a heart-shaped (bicornate)
uterus, which makes it difficult to carry a pregnancy. People with WAGR syndrome have an
increased risk (estimated at 45 percent) of developing Wilms tumor, a rare form of kidney
cancer. This type of cancer is most often diagnosed in children, but is sometimes seen in
adults. Other signs and symptoms of WAGR syndrome can include childhood-onset obesity,
inflammation of the pancreas (pancreatitis), and kidney failure. When WAGR syndrome
includes childhood-onset obesity, it is often referred to as WAGRO syndrome. The prevalence
of WAGR syndrome is unknown; however, it appears to be a rare condition. It is estimated
that one-third of people with aniridia actually have WAGR syndrome. Approximately 7 in 1,000
cases of Wilms tumor can be attributed to WAGR syndrome. WAGR syndrome is caused by a
deletion of genetic material on the short (p) arm of chromosome 11. The size of the deletion
varies among affected individuals. The signs and symptoms of WAGR syndrome are related
to the loss of multiple genes on the short arm of chromosome 11. WAGR syndrome is often
described as a contiguous gene deletion syndrome because it results from the loss of several
neighboring genes. The PAX6 and WT1 genes are always deleted in people with the typical
signs and symptoms of this disorder. These genes play significant roles in early development.
Researchers believe that loss of the PAX6 gene is associated with the characteristic eye
features of WAGR syndrome and may also affect brain development. Deletion of the WT1
gene appears to be associated with Wilms tumor and the genitourinary abnormalities.
Researchers are working to identify additional genes deleted in people with WAGR syndrome
and determine how their loss leads to the other features of the disorder. Most cases of WAGR
syndrome are not inherited. They result from a chromosomal deletion that occurs as a random
event during the formation of reproductive cells (eggs or sperm) or in early fetal development.
Affected people typically have no history of the disorder in their family. Some affected
individuals inherit a chromosome 11 with a deleted segment from an unaffected parent. In
these cases, the parent carries a chromosomal rearrangement called a balanced
translocation, in which no genetic material is gained or lost. Balanced translocations usually
do not cause any health problems; however, they can become unbalanced as they are passed
to the next generation. Children who inherit an unbalanced translocation can have a
chromosomal rearrangement with extra or missing genetic material. Individuals with WAGR
syndrome who inherit an unbalanced translocation are missing genetic material from the short
arm of chromosome 11, which results in an increased risk for Wilms tumor, aniridia,
genitourinary anomalies, and intellectual disability characteristic of this disorder.
Sebenta de Gentica
FCML
180
Deleco no cromossoma 13 e retinoblastoma
(Ver pgina 166)
Citogentica de clulas neoplsicas
It has long been appreciated that tumor cells carry chromosomal aberrations. Errors in the
faithful segregation of chromosomes during mitotic or meiotic cell division play a crucial role in
the generation of chromosomal aneuploidies, the genetic consequences of which are
increased copy numbers of oncogenes and loss of tumor suppressor genes. Cytogenetic
abnormalities that affect the integrity of a chromosome are referred to as structural
aberrations. These can be either balanced or unbalanced translocations (ver pgina 122). In
particular, balanced chromosomal translocations are frequently observed and considered
pathogenetic events in haematopoietic malignancies.
One outcome of a translocation is the re-positioning of an oncogene in proximity to a strong
enhancer from another gene. This results in overexpression of the oncogene and can lead to
cellular transformation. In human Burkitt's Lymphoma, mouse plasmacytomas and pro-B cell
lymphomas, the c-myc oncogene is juxtaposed to the enhancer for the immunoglobulin heavy
chain gene. Dysregulation of c-myc in this manner is believed to increase both the rate of cell
division and chromosome instability. Balanced chromosomal translocations can have
oncogenic effects through the production of fusion proteins. For instance, in almost 95% of
chronic myelogenous leukemia (CML) case a translocation between chromosomes 9 and 22
results in the formation of what is commonly referred to as the Philadelphia chromosome. This
results in the fusion of the BCR and ABL genes. The BCR-ABL fusion protein has increased
tyrosine kinase activity and transforms haematopoietic cells. Inhibition of the constitutive
kinase activity with the BCR-ABL-specific inhibitor STI571 is currently in clinical trials and the
overall response rate of patients in blast crisis in one study was reported to be 55%, with
complete remission in 11% of the patients.
Karyotype analysis based on G- or R-banding techniques have been widely applied to the
characterization of cytogenetic abnormalities in tumor cells and have contributed significantly
to the identification of recurrently involved chromosomal loci and hence to the molecular
cloning of cancer causing genes. In many instances, however, the cytogenetic analysis of
chromosomes from solid tumors has proven to be challenging. This is due to the often-low
mitotic index, the poor quality of metaphase chromosomes, and the sheer number of
cytogenetic abnormalities. The use of genetically engineered mice as model systems of
human cancer has fueled the need for better cytogenetic analysis in this species as well.
Mouse chromosomes do not vary much in size and all are acrocentric. Therefore, some of the
Sebenta de Gentica
FCML
181
parameters used for identification of human chromosomes are less helpful, making
karyotyping very difficult. A further drawback at present is that rather few cytogeneticists are
trained in karyotyping mouse chromosomes.
The ability to microscopically visualize the location of individual sequences on metaphase
chromosomes using fluorescence in situ hybridization (FISH) techniques has greatly
broadened the application of cytogenetic methodologies, ushering in the era of molecular
cytogenetics (for historical review. FISH probes come in many varieties. They can be specific
to (A) unique regions and genes, (B) repetitive sequences such as telomeres and
centromeres, (C) entire chromosomes, chromosome bands or arms and (D) in the case of
comparative genomic hybridization (CGH) as large as entire genomes. Advances in
fluorochrome chemistry (i.e. stability, stoke shift, quantum yield, and conjugability to
nucleotides) and photodetection devices (i.e., charged coupled device cameras and
photomultiplier tubes) have further enhanced the sensitivity and multiplicity of FISH. The
merging of these technical advances is reflected in the combinatorial labeling and
hybridization techniques known as comparative genomic hybridization (CGH), spectral
karyotyping (SKY), and m-FISH. CGH is a quantitative method for comparing the copy
number of genomic regions between a control (DNA extracted from karyotypically normal
cells) and test sample (i.e. tumor DNA). SKY and m-FISH involve the hybridization of
differentially labeled chromosome painting probes for the identification of both numerical and
structural aberrations in the genome. Each of these two conceptual methodologies alone
provides a significant advance in the way we analyze genomes. Together, SKY, m-FISH and
CGH offer novel tools to understand tumor specific and stage specific chromosome
aberrations.
Spectral Karyotyping
Spectral karyotyping (SKY) is a molecular cytogenetic technique used to generate a color-
coded karyotype of human and mouse chromosomes. Prior to its development,
cytogeneticists analyzed tumors and leukemias using a Giemsa staining technique that
produced a dark and light banding pattern on chromosomes (caritipo). As previously
mentioned, chromosomal analysis using this technique alone can be challenging since tumor
metaphase chromosomes are often poorly spread, have many rearrangements and subtle
translocations or insertions. In combination with chromosome banding techniques, SKY allows
for a more comprehensive description of cytogenetic abnormalities. SKY has greatly
enhanced the ability to analyze complex karyotypes from both human tumors and murine
model systems.
Sebenta de Gentica
FCML
182
Fig. 87: Schematic representation of the steps involved in SKY analysis
Comparative Genomic Hybridization- CGH
CGH employs two-color fluorescence in situ hybridization of both tumor DNA and normal
control DNA to normal metaphase chromosomes. This molecular cytogenetic technique
measures the genetic imbalances within tumor genomes, and has become an exceedingly
valuable tool for the analysis of chromosomal aberrations in solid tumors. One major
advantage of CGH is that only genomic tumor DNA is needed for analysis. Therefore, DNA
extracted from archived, fixed tumors as well as from microdissected tissue can be analyzed.
After extraction of test DNA and normal DNA, the samples are differentially labeled with
discernable fluorochromes (i.e. tumor DNA with FITC [green] and control DNA with TRITC
[red]) (Fig. 72 3A). The 2genomes are combined with an excess of human Cot-1 DNA and
then hybridized to normal metaphase chromosomes (Fig. 72 3B). Images of metaphase
spreads are then acquired with a CCD camera and fluorochrome-specific optical filter sets to
Sebenta de Gentica
FCML
183
capture the FITC and TRITC fluorescence (Fig. 72 3C). Differences in fluorescence intensity
values between tumor and control DNA represent gains and losses of specific chromosomes
or chromosomal regions. For example, a gain of a chromosomal region in the test sample
would result in an increased intensity of green fluorescence. A loss within a chromosomal
region in the tumor would be indicated by a shift towards red intensities. Specialized CGH
analysis software measures fluorescence intensity values along the length of the
chromosomes and translates the ratios into chromosome profiles. The ratio of green to red
fluorescence values is used to quantitate genetic imbalances in tumor samples. Further
modification of the CGH technique includes the replacement of metaphase chromosomes with
unique DNA sequences spotted in arrays on a glass slide. Fluorescence intensities in both
test and reference DNA hybridizations to the immobilized sequences on the array (i.e. cDNA,
BAC or oligos) are averaged and normalized, and can be used to calculate an increase or
decrease in copy number. This array CGH allows for higher resolution of closely spaced
genomic aberrations as well as the detection of microdeletions
Fig. 88: Schematic representation of Comparative Genomic Hybridization
Sebenta de Gentica
FCML
184
Comparative Cytogenetics
Cytogenetic analysis of human tumors reveals a non-random distribution of chromosome
rearrangements resulting in genomic imbalances. Such aberrations are tissue specific and in
some cases are fingerprints for a specific tumor. To better understand the significance of
these chromosomal aberrations and identify in a controlled setting those that are early and
disease initiating events, animal models of human cancer are extremely beneficial. The ability
to generate gene-specific null mutants in mice affords us the opportunity to elucidate
biochemical pathways leading to tumorigenesis and to query the relationship between
chromosomal aberrations and the consequences of gene mutation on genomic stability. In
addition, the generation of mouse strains hetero- or homozygously deficient for individual
genes, the use of conditional knockouts, and inducible systems for transiently repressing or
activating target genes increases the value of mouse models, particularly for cancer. Because
the cytogenetic abnormalities observed in human cancers are fundamental, and often
distinguishing features, it is extremely useful to evaluate whether the patterns of genomic
imbalances are similar in human tumors and their corresponding mouse models. The
application of molecular cytogenetic techniques, such as SKY and CGH, has been a key
factor in identifying similarities between human diseases and their respective mouse models.
Mammary cancer is just one such example. The construction of human-mouse orthology
maps and the sequencing of both genomes has greatly aided the process of comparative
cytogenetics. For example, humans have 22 autosome pairs that vary greatly in size and most
have two visible arms (p & q) separated by a centromere. Mice, however, have 19 autosome
pairs of relatively similar size with one arm and a centromere near the end. Specific
chromosomes or chromosome regions of one species can be mapped onto the chromosomes
of different species, making it possible to follow chromosome reshuffling during evolution (fig.
73). For instance, mouse chromosome 11 contains sequences orthologous to human
chromosome 17, as well as regions of synteny with human 2p (red), 5q (gold), 7p (light pink),
16p (orange) and 22q (dark pink) (fig. 74). By following this comparative cytogenetic
approach, we can evaluate chromosome gains and losses in similar tumor types from different
species to determine if the same set of oncogenes or tumor suppressor genes is amplified or
lost, respectively. Because genomic imbalances, generated primarily through chromosomal
translocations or aneuploidies, are specific to each type of human malignancy, comparative
analyses provides a validation of murine models and an entry point for the identification of
new genes involved in tumorigenesis.
Sebenta de Gentica
FCML
185
Fig. 89: Orthology map of human and mouse chromosomes
Fig. 90: Synteny between mouse chromosome 11 (MMU11) and human chromosome 17(HSA17).
Cytogenetic Analysis of Mouse Models of Haematopoietic Malignancies
Haematopoietic malignancies are quite distinct from solid tumors in that rather than gross
chromosomal aneuploidy, the causative genomic alterations tend to result from the specific
juxtaposition of genes or gene segments at the site of translocations. We have analyzed pro-B
cell lymphomas, plasmacytomas and thymic lymphomas from a variety of different mouse
models. In mice deficient for the gene mutated in ataxia telangiectasia (ATM) thymic
lymphomas develop in nearly all of the animals between 2 and 4 months of age. These tumors
exhibit a variety of chromosomal aberrations, including translocations, insertions, deletions,
and duplications. Through the use of locus-specific BAC clones, we were able to determine
Sebenta de Gentica
FCML
186
that the breakpoint on chromosome 14 occurs at the T-cell receptor (Tcr) locus during the
programmed genomic recombination process known as V(D)J recombination. Other common
aberrations included a deletion of the distal portion of chromosome 12 resulting in loss of the
immunoglobulin heavy chain gene (IgH) as well as trisomy of chromosome 15, which contains
the c-myc oncogene. Aberrations involving the T-cell receptor locus are also found in human
ataxia telangiectasia patients and support the use of the Atm mouse as a model for
tumorigenesis in the absence of this gene product. Mice deficient in Ku80, a protein involved
in the non-homologous end joining (NHEJ) repair of DNA double strand breaks, also develop
thymic lymphomas, but at a low frequency. These tumors almost invariably contain extra
copies of chromosome 15. When these, and mice deficient in other NHEJ proteins, are
crossed onto a p53-deficient background, 100% of the mice now develop tumors, the
spectrum of which shifts from T-cell to pro-B cell lymphoma. These tumors all harbor
translocations involving chromosome 12 and 15 resulting in the juxtaposition and co-
amplification of the IgH enhancer and the c-myc oncogene. While this rearrangement on the
surface resembled that seen in human Burkitt's lymphoma, whereby the IgH enhancer and c-
myc are also brought into close proximity, closer examination of the breakpoints in the mice
revealed that the mechanism of translocation in these animals is quite distinct from that
observed in the human disease and more closely reflects the type of aberrations (i.e. non-
reciprocal translocations, gene amplification and complex chromosome rearrangements) often
seen in the solid tumors. Another mouse model, which perhaps more closely mimics human
Burkitt's lymphoma or multiple myeloma, is the induction of plasmacytomas in specific mouse
strains. In this system the rearrangements are believed to occur primarily in mature B-cells
during the activation induced deaminase (AID)-dependent process of class switch
recombination, not during V(D)J-mediated antigen receptor rearrangement in developing B-
cells. The rearrangements involving the IgH locus on chromosome 12 and the c-myc locus on
chromosome 15 are typically reciprocal in nature. However, in some instances amplification of
the juxtaposed IgH & c-myc loci have been observed. Additionally, other differences are
evident between this mouse model and the human diseases. Often in human haematopoietic
malignancies, translocations result in the joining of two different genes and the production of
fusion proteins. Mouse models have been developed in which the fusion protein is introduced
as a transgene. Such mice are extremely useful for studying the effects of expression of these
chimeric molecules during the earliest stages of tumorigenesis. Identification of functional
elements within the fusion proteins has been achieved through the introduction of transgenes
in which various regions have been modified. Thus, the use of mouse models for the study of
human haematopoietic malignancies has proven to be extremely fruitful.
Sebenta de Gentica
FCML
187
Cytogenetic Analysis of Mouse Models of Breast Cancer
Human breast carcinomas are defined by copy number increases frequently mapping to
chromosomes 1q, 8q, 17q and 20q where a variety of oncogenes involved in breast cancer
have been identified. With this knowledge in hand, we applied SKY and CGH to the analysis
of primary mammary tumors arising in knock-out mice of the tumor suppressor gene Brca1,
mice transgenic for the oncogenes c-Myc or HER2/Neu, and mice expressing the polyoma
virus middle T antigen or the SV40 large T antigen under control of the mammary gland tissue
specific promoter MMTV. The distribution of gains and losses in each system revealed
recurrent patterns of chromosomal aberrations, although each model was quite distinct. The
Brca1 KO, however, contained aberrations more consistent with the rearrangements observed
in human breast carcinomas. The major imbalance observed in 58% of the HER2/Neu
transgenic mouse tumors was deletion of chromosome 4 (MMU4) bands C-E. This suggested
that in addition to HER2/Neu overexpression, decreased expression of a gene(s) in this region
of chromosome 4 was important for the induction of mammary gland transformation (Figure
6). The distal portion of mouse chromosome 4 is orthologous to human 1p31-36 and 9p21, the
latter of which contains the INK4 locus. In both the HER2/Neu and the C3(1)SV40Tag models,
oncogene amplification results from the presence of acentric extra chromosomal fragments
known as double minute chromosomes (dmin). Dmin play a critical role in tumor cell genetics
where they are frequently associated with the overexpression of oncogene products. FISH
using locus specific probes revealed that amplification of the HER2/Neu and K-Ras
oncogenes was manifested as dmin in the HER2/Neu and C3(1)SV40Tag mouse models,
respectively (Figure 7). This finding suggests that in addition to the presence of a strong initial
stimulus for cellular transformation (i.e. the overexpression of an oncogene or the presence of
a viral protein), secondary genetic alterations are necessary for tumor formation, at least in the
mammary gland. These results are consistent with the proposed step-wise increase in
chromosome aberrations that parallels tumorigenic progression in human colorectal and
cervical carcinomas.
Sebenta de Gentica
FCML
188
Fig.91: Chromosome gains and losses in the Her2/Neu transgenic mouse model.
A comparison of chromosomal alterations between mouse models and their human tumor
counterparts enables further refinement of those regions necessary for tumorigenesis. For
example, the region most commonly gained in mammary tumors is the distal portion of
chromosome 11. This region is orthologous to an area of the human genome (17q25)
amplified in human breast cancer as well as other types of epithelial tumors. The tumor
suppressor genes BRCA1 and Trp53, as well as the oncogenes HER2/Neu, Tbx2, Rad51c
and Grb2 are located in MMU11C-E and human 17q25. However, because the oncogenes in
the mouse map closer to the centromere and are outside the region of amplification, we
conclude that they are not responsible for tumorigenesis in the mouse, and therefore unlikely
to be involved in human breast cancer development. Thus, another candidate gene(s) with
oncogenic potential residing on human chromosome 17 may be important for mammary gland
transformation.
Cytogenetic experiments conducted in mice with the use of comparative maps are an
important tool for understanding the sequence of genetic events required for tumorigenesis.
The results obtained can be used to better understand the processes involved in cellular
transformation and for the identification of molecular pathways conserved between mice and
humans. Increasing the cytogenetic resolution is extremely useful not only for describing
genetic alterations, but also for defining those genomic changes that are specific to each
tumor type. This has largely been made possible through the combined application of CGH
(for identifying regions of gain and loss) and SKY (to identify the mechanism by which the
regional copy number or gene expression is altered). Further sub-classification of tumors into
Sebenta de Gentica
FCML
189
more specific categories based on their genomic profile may prove useful in tailoring or
identifying responsiveness to particular therapies.
Actividade enzimtica da telomerase e o estudo de neoplasias
Telomerase, a eukaryotic ribonucleoprotein (RNP) complex, contains
both an essential RNA
and a protein reverse transcriptase subunit.
By reverse transcription, the telomerase RNP
maintains telomere
length stability in almost all cancer cells. Over the past few
years there has
been significant progress in identifying the
components of the telomerase holoenzyme
complex and the proteins
that associate with telomeres, in order to elucidate mechanisms
of
telomere length regulation. This review covers recent advances
in the field including the use of
telomerase in cancer diagnostics
and an overview of anti-telomerase cancer therapeutic
approaches.
A fundamental difference in the behavior of normal versus tumor
cells in culture is
that normal cells divide for
a limited number of times (exhibit cellular senescence) whereas
tumor cells usually have the ability to proliferate indefinitely
(are immortal). There is substantial
experimental evidence that
cellular aging is dependent on cell division and that the total
cellular lifespan is measured by the number of cell generations,
not by chronological time. This
means there is an intrinsic
molecular counting process occurring during cell growth that
culminates in the cessation of cell division. It is now evident
that the progressive loss of the
telomeric ends of chromosomes
is an important timing mechanism in human cellular aging.
Human telomeres contain long stretches of the repetitive sequence
TTAGGG which are bound
by specific proteins. With each
cell division, telomeres shorten by 50200 bp, primarily
because the lagging strand of DNA synthesis is unable to replicate
the extreme 3' end of the
chromosome (known as the end replication
problem). When telomeres become sufficiently
short,
cells enter an irreversible growth arrest called cellular senescence.
In most instances
cells become senescent before they can accumulate
enough mutations to become cancerous,
thus the growth arrest
induced by short telomeres may be a potent anti-cancer mechanism.
Telomerase, a eukaryotic ribonucleoprotein (RNP) complex,
helps to stabilize telomere length
in human stem cells, reproductive
cells and cancer cells by adding TTAGGG repeats
onto the
telomeres using its intrinsic RNA as a template for
reverse transcription. Telomerase activity
has been found
in almost all human tumors but not in adjacent normal cells
.
The most
prominent hypothesis is that maintenance of
telomere stability is required for the long-term
proliferation
of tumors. Thus, escape from cellular senescence
and becoming immortal by
activating telomerase, or an alternative
mechanism to maintain telomeres, constitutes an
additional
step in oncogenesis that most tumors require for their ongoing
proliferation. This
makes telomerase a target not only for cancer
diagnosis but also for the development of novel
anti-cancer
therapeutic agents.
The telomerase RNP and telomere complex present multiple
Sebenta de Gentica
FCML
190
potential
targets for the design of new anticancer strategies. Telomerase may be a challenging
target since its inhibition
should exhibit a lag phase: the lack of telomerase should not
affect
cell growth rates until progressive telomere shortening
with each cell division eventually
causes cells to die or undergo
growth arrest. Although it has been correctly suggested that
this
approach would not be sufficient by itself in patients
with a large tumor burden, it may be a
unique
approach to patients with minimal residual disease. Importantly,
normal somatic cells
that lack telomerase expression should
be largely unaffected by anti-telomerase therapy.
Although telomerase
inhibitors should possess great specificity, it is hoped they
will also
display low toxicity and few side effects. The most
likely use of telomerase inhibitors would be
as an adjuvant
treatment in combination with surgery, radiation treatment and
typical
chemotherapy, when tumor burden is minimal. It is also
possible that telomerase inhibitors
could be used following
standard therapies in which there is no clinical evidence of
residual
disease in order to treat possible micrometastases,
and thus prevent cancer relapse. These
situations will require
prolonged treatment, so it will be important that the drugs
have a low
toxicity profile and are easily administered.
Para alm da telomerase, existe outro processo atravs do qual os tumores conseguem
fazer de forma alternativa a manuteno dos telmeros: o ALT.
No ALT, uma cadeia de DNA de um telmero liga-se a uma cadeia complementar de outro
telmero, obtendo assim o modelo para a sntese do novo DNA telomrico. Abrem-se as
duas cadeias como se v no esquema seguinte, servindo uma de molde outra. Aps a crise
celular, alguns telmeros ficam to curtos que as repeties telomricas encontram-se logo
aps a zona subtelomrica do cromossoma, onde j no existem as repeties da sequncia
telomrica. Quando se d o mecanismo ALT, possveis sequncias GGTTAG perdidas nas
zonas subtelomricas podem agir como primer de adio de novo DNA telomrico,
resultando isto na substituio das variaes subtelomricas por repeties telomricas
GGTTAG. Os telmeros mantidos pelo ALT apresentam elevada heterogeneidade e so
observados em clulas que tambm apresentam telomerase.
Oncobiologia:
Sebenta de Gentica
FCML
191
Sebenta de Gentica
FCML
192
A GENTICA MOLECULAR HUMANA - REAS DE INTERVENO
1. Testes genticos predictivos_________________________________
Diagnstico pr-implantatrio
O PGD um mtodo usado para determinar doenas genticas em embries obtidos por
fertilizao in- vitro (IVF), antes do seu desenvolvimento. A IVF consiste em remover um
vulo da mulher que posteriormente fertilizado com esperma do dador, sendo que este
embrio apenas implantado no tero se nenhuma doena gentica for detectada.
essencialmente procurado por progenitores portadores de doenas geneticamente
transmissveis de forma a evitar a transmisso aos filhos. Consiste em remover uma ou duas
clulas do embrio, sendo este mantido fixo numa posio com a ajuda de uma pipeta
enquanto outra o perfura e remove as clulas por ligeira suco. O DNA ento isolado e
amplificado por PCR, procedendo-se sua anlise molecular. Quando algum defeito gentico
detectado procede-se destruio desse embrio, caso contrrio este ento implantado
no tero. Relativamente fase do embrio em que estas clulas so removidas, varia entre o
blastmero e o blastocisto. O primeiro forma-se 3 dias aps a IVF e formado por 4 a 10
clulas, sendo estas totipotentes (define-se como totipotente a capacidade que uma nica
clula removida tem de originar ela sozinha o embrio) ; o segundo forma-se 5 a 6 dias aps
a IVF. Devido totipotncia das clulas do blastmero, pases como a Alemanha defendem a
realizao do PGD apenas no estdio de blastocisto j que proibida a destruio de clulas
humanas totipotentes. O PGD permite a deteco de um grande nmero de doenas entre as
Sebenta de Gentica
FCML
193
quais a trissomia 21 e outras aneuplodias cromossomais, sendo essencialmente vantajosa
em casos de sucessivos abortos espontneos, insucesso na IVF e idade avanada da
mulher. Permite detectar doenas associadas ao cromossoma X, como a distrofia muscular
de Duchenne, e outras doenas como a atrofia da espinal medula tipo I, a doena de
Huntigton e a fibrose qustica. Este processo tem como vantagens permitir que pais
portadores de doenas geneticamente transmissveis assegurem o nascimento de crianas
saudveis, reduzir a incidncia de um grande nmero de doenas, assim como diminuir os
custos nos sistemas mdicos j que o tratamento de grande parte destas altamente
dispendioso. Comparativamente amniocintese (processo que tambm permite a deteco
de doenas genticas), o PGD no apresenta riscos para a me e evita a realizao do
aborto, necessrio efectuar quando a doena detectada por anlise do lquido amnitico.
Contudo, s aplicvel quando efectuada IVF, no o sendo numa gravidez pelo mtodo
natural, e levanta grandes questes de tica porque h quem defenda que, apesar de com
apenas 5 ou 6 dias, o embrio j uma vida humana, e como tal, opem-se sua destruio.
Diagnstico pr-natal
Diagnstico Pr Natal o conjunto de procedimentos que permitem detectar determinadas
anomalias congnitas antes do nascimento. A primeira fase do desenvolvimento pr-natal de
um ser humano compreende a diviso, a implantao e a primeira diferenciao em duas
camadas distintas de clulas, momento em que se regista uma percentagem considervel de
abortos espontneos. A segunda corresponde fase embrionria e quando ocorrem as
maiores anomalias morfolgicas. A ltima a fase em que ocorrem os defeitos fisiolgicos e
as menores anomalias morfolgicas.
No despiste de mutaes cromossmicas, sejam elas anomalias estruturais ou numricas (de
origem familiar ou de novo), importante uma anlise citogentica das clulas na fase fetal.
Esta anlise pode ser feita utilizando trs tipos de material e a escolha deste material
depende de cada caso. A primeira anlise, conhecida como amniocentese, baseia-se na
recolha de lquido amnitico, que contm clulas da pele do feto, e realizado sob controlo
ecogrfico. Pode ser feito a partir da 13 semana de gravidez e o exame que apresenta
menores riscos de aborto (inferior a um por cento) j que no to invasivo. A desvantagem
deste mtodo a demora na obteno dos resultados, que pode ir de duas a quatro
semanas. O segundo exame analisa as clulas das vilosidades do crion (parte da placenta
que vem do feto), mais invasivo, mas tem a vantagem de poder ser realizado numa fase
muito precoce do desenvolvimento do feto (9/12 semanas) e de se obterem resultados em
24/48 horas. A cordocentese o terceiro exame e utiliza como material o sangue fetal
(extrado do cordo umbilical). feito mais tarde, a partir da 20 semana de gravidez. Os dois
Sebenta de Gentica
FCML
194
ltimos so os mais invasivos e com maior risco de aborto. De qualquer das formas, o mdico
que acompanha a gravidez saber se h necessidade de efectuar estes testes e qual deles,
tendo em considerao determinados factores importantes. O DPN aconselhado em
diversos casos, um deles a idade materna avanada. Verifica-se uma maior incidncia de
anomalias nos fetos de mes com mais de 45 anos (em mdia, um em cada 25
nascimentos), enquanto que nas mes com menos de 30 anos os valores de um em cada
1.500. aconselhado, tambm, nos casos em que um dos progenitores possui alguma
anomalia cromossmica ou se o casal tem um filho nessas circunstncias. No caso de, numa
ecografia, se detectar alguma malformao congnita ou de haver ambiguidade sexual
importante esta anlise j que pode sugerir uma anomalia similar. De considerar ainda casais
com histria de infertilidade, abortos espontneos ou neoplasias (leucemias, tumores).
Diagnstico pr-sintomtico
possvel, desde 1995, proceder ao diagnstico pr-sintomtico da doena, de uma forma
segura e fivel. Esta tcnica permite ter conhecimento sobre se o indivduo ou no portador
de doenas, antes da manifestao de quaisquer sintomas, graas clonagem do gene da
doena e identificao da respectiva mutao. Os aspectos psicolgicos so importantes
para o aconselhamento gentico e para a interveno nas doenas neurolgicas de
aparecimento tardio (como por exemplo a doena de Huntington, doena de Machado Joseph
e polineuropatia amiloidtica familiar)
Aconselhamento gentico, avaliao de risco e preveno da recorrncia
O conhecimento cientfico no campo da gentica humada pode ser usado no aconselhamento
gentico de famlias em que h suspeitas de um risco aumentado para anomalias ou doenas
hereditrias em filhos futuros. O aconselhamento gentico refere-se totalidade das
actividades que:
- Estabelecem o diagnstico dessas doenas
- Avaliam o risco de recorrncia
- Comunicam ao paciente e sua famlia a probabilidade de recurrncia
- Fornecem informao e aconselhamento relativamente aos diversos problemas
levantados pela doena e pela sua histria natural, incluindo os potenciais fardos a nvel
mdico, econmico, psicolgico e social
- Fornecem informao relativamente s opes a tomar a nvel reprodutivo, incluindo o
diagnstico pr-natal
- Encaminham os pacientes para os especialistas indicados
Sebenta de Gentica
FCML
195
O mdico tem a responsabilidade de fornecer um aconselhamento gentico adequado ao
indivduo que apresente um determinado problema gentico. Este aconselhamento deve ser
feito em caso de doena conhecida ou suspeita, no caso de portadores, no diagnstico pr-
natal em casos de consanguinidade exposio a um agente teratognico ou quando existe
histria de abortamentos anteriores. Esta aco tem como objectivo prevenir os pais para
problemas que possam surgir nos filhos, para prevenir a doena com uma teraputica
adequada (se existir), promover um meio ambiente adequado ao doente e dar um apoio
psicolgico nos casos que tal o indiquem. O aconselhamento gentico uma actividade
medicamente orientada e no eugnica. Os casais que procuram aconselhamento so
encorajados a tomar as suas prprias decises face reproduo segundo o que acharem
mais adequado, independentemente dos possveis efeitos nocivos no pool gnico da
populao. Esta prtica coloca o aconselhamento dentro dos parmetros da prtica mdica,
em que o indivduo e a sua famlia, e no a populao em geral, o foco de aconselhamento
e tratamento. A avaliao do risco e a preveno da ocorrncia so problemas fundamentais
no aconselhamento gentico, uma vez que permitem, tendo em conta os princpios de
transmisso de caracteres aos descendentes, desaconselhar ou no uma gravidez ou
pronunciar-se sobre a possvel interrupo da mesma. O risco gentico em doenas
mendelianas est claramente definido e depende do modo de hereditariedade especfico. Os
riscos clnicos reais para o paciente, nomeadamente em doenas autossmicas dominantes,
dependem tambm da expressividade e penetrncia variveis e ainda da manifestao tardia
de muitas das doenas. s vezes possvel alterar as estimativas do risco mendeliano
usando a anlise de Bayes (mtodo de avaliao da probabilidade relativa de duas
possibilidades opcionais, em geral as de um indivduo possuir ou no um dado alelo) aplicada
ao heredograma, que leva em conta informaes sobre a famlia que podem elevar ou reduzir
o risco mendeliano anterior. Quando no se dispe de nmeros de risco mendeliano, deve-se
confiar nos valores do risco emprico, que permitem apenas fazer uma estimativa com base
em experincias prvias. Para muitas famlias que procuram aconselhamento, um dos
objectivos principais evitar a recorrncia do distrbio especfico. As medidas empregues
nesse sentido so:
- Exames laboratoriais genticos
- Contracepo ou esterilizao
- Adopo
- Inseminao artificial com smen de dador, adequada a situaes em que o futuro pai
portador de um gene para uma doena autossmica dominante ou ligada ao X (ou da
inseminao in-vitro do vulo por um espermatozode Y, no caso da doena associada ao
X)
Sebenta de Gentica
FCML
196
- Anlise do DNA de embries em estdios pr-implantatrios recorrendo ao mtodo PCR
- Diagnstico pr-natal e interrupo da gravidez, se desejado e face a um resultado positivo.
2. Rastreios e diagnstico precoce (fenilcetonuria e hipotiridismo)
Teste do pezinho!
3. A Medicina Molecular
A farmacogenmica e a oncogenmica
A farmacogentica uma rea especial da gentica bioqumica que lida com as variaes na
resposta a frmacos e a contribuio da gentica para tais variaes. Um exemplo clssico
o efeito dos barbitricos no desencadeamento do ataque de porfiria em indivduos com o
gene para a porfiria aguda intermitente. Tendo em conta a variao normal na resposta a
frmacos, define-se potncia de um medicamento, como a dose que produz um dado efeito
em pelo menos 50% da populao. Quando se administra uma dose standard de um
determinado frmaco a um nmero elevado de indivduos normais de uma populao,
observam-se variaes considerveis dos nveis sricos desse frmaco entre diferentes
indivduos. Embora existam numerosos factores que podem influenciar esses nveis, os seus
principais determinantes so variao nas taxas de metabolismo, determinadas pela semi-
vida do frmaco. A distribuio destes nveis ajusta-se normalmente a uma curva de Gauss e
um certo nmero de pessoas, em cada extremidade dessa curva apresenta, ou nveis
demasiado baixos ou demasiado altos aps a administrao de uma dose mdia. O resultado
, quer toxicidade, quer ausncia de efeito teraputico. No entanto, a resposta a frmacos
pode exibir uma distribuio descontnua com distines ntidas entre graus diferentes de
resposta. Uma distribuio populacional, bi ou tri-modal, da actividade de uma enzima
implicada no metabolismo de um frmaco pode indicar que a enzima codificada por genes
num locus polimorfo. Uma outra situao que tambm objecto de estudo por parte da
farmacogentica so as interaces medicamentosas, em que a administrao simultnea de
dois frmacos leva reduo ou aumento dos nveis de pelo menos um deles (por exemplo,
devido ao facto de um dos frmacos induzir o aumento da actividade dos sistemas
enzimticos de metabolismo de xenobiticos).
Protemica:
A protemica a anlise sistemtica de todas as sequncias proteicas e padres de
expresso das protenas nos tecidos, que envolvem o isolamento, a separao, a
identificao e caracterizao funcional de todas as protenas num organismo.
Caractersticas que a distinguem da transcriptmica:
Sebenta de Gentica
FCML
197
- existncia de RNAm que no so traduzidos
- turnover proteco independente da abundncia de transcriptos
- modificaes pstraduo
Protemica de expresso: comparao entre amostras distintas: expresso proteca de
um tumor com metstases vs sem metstases; com resposta vs sem resposta
quimioterapia, etc
Tcnicas:
a) Electroforese bi-dimensional(2DGE)
b) Espectrometria de massa (EM)- digesto peptdica com tripsina
Protemica de interao:
- definio de uma rede de interao entre protenas
- estrutura 3Dproteca
Tcnicas:
a) Sistema 2-hbridos
- Factores de transcrio tem 2 domnios funcionais independentes: domnio de ligao ao
DNA (BD) e domnio de activao de transcrio (AD)
- A interao entre 2 protenas traduz a presena de um factor de transcrio
funcionalmente activo
NOTA: A protemica tender a substituir a transcriptmica na definio do diagnstico e
prognstico em patologia humana. A protemica premitir identificar novas vias moleculares
em patologia humana, particularmente no cancro.
Southern Blot:
Esta uma tcnica utilizada frequentamente na anlise de genes. Consiste da clivagem da
totalidade do DNA humano, por enzimas de restrio, em milhares de fragmentos. Estes so
separados de acordo com o seu peso molecular atravs de uma electroforese em gel de
agarose, sendo as duplas hlices separadas em seguida por um tratamento com substncias
alcalinas, de modo a produzir fragmentos de DNA monocatenrio. Os fragmentos so ento
blotted num filtro de nitrocelulose e fixados a 80C. O padro resultante representa uma
rplica dos fragmentos de DNA separados electroforeticamente no gel de agarose original.
Estes podem ser identificados por hibridao com sondas de DNA marcadas
radioactivamente e especficas para genes ou regies cromossmicas. Qualquer fragmento
Sebenta de Gentica
FCML
198
que contenha parte ou a totalidade da sequncia sondada ser visvel como uma banda
escura aps autorradiografia.
Este procedimento til na determinao do nmero de cpias de determinado gene
existentes em determinado tecido ou na identificao de alteraes grosseiras no gene, como
deleces, inseres ou inverses. Ocasionalmente, se uma base especfica alterada ao
nvel do prprio local de restrio, esta tcnica pode detectar uma point mutation.
Enzimas de restrio: so endonucleases que reconhecem sequncias consensuais de
DNA.
Clonagem:
Clonagem funcional: doena funo gene mapeamento
Clonagem aposicional: doena mapeamento gene funo
Procedimentos para a clonagem:
Para clonar um segmento do genoma necessrio ter uma sequncia de DNA bicatenar.
Quando se parte de RNA, este convertido em cDNA, mediante uma transcriptase inversa. O
primeiro passo para clonar DNA por meio de recombinao gentica consiste na sua
fragmentao por uma enzima de restrio seleccionada em funo do tamanho dos
fragmentos que se deseja obter. Estas enzimas so produzidas por algumas bactrias para
clivarem o DNA estranho e, dessa forma, se protegerem da invaso por microorganismos,
nomeadamente bacterifagos. As enzimas de restrio fragmentam o DNA sempre que
reconhecem determinadas sequncias especficas que variam habitualmente entre 4 e 8
bases.
A enzima e o vector devem ser seleccionados em conjunto, pois o vector deve possuir a
sequncia de restrio reconhecida pela mesma enzima que usada para fragmentar o DNA.
H diferentes vectores disponveis para clonar DNA, com capacidades diferentes, de modo a
poderem inserir fragmentos de DNA de tamanho diverso. Os plasmdeos so usados para
clonar fragmentos de DNA com um tamanho mdio de 4kb, os fagos at 20kb e os
cosmdeos at 40kb. Fragmentos maiores, at cerca de 350kb, podem ser inseridos em
BACs (bacterial artificial chromossomes). Para fragmentos ainda maiores, podendo atingir 1,5
Mb, recorre-se a YACs (yeast artificial chromossomes).
Sebenta de Gentica
FCML
199
Random:
- Terapia gnica resulta melhor em doenas autossmicas recessivas.
- Hereditabilidade uma medida de proporo da variao devida a causas genticas
- BLAST (Basic Local Alignment Search Tool) is a family of programs used in genetics
research, which try to identify similarities between a given sequence of amino acids or
nucleotides, and a database containing proteins or DNAs. The blast programs compare a
given sequence (of amino acids or nucleotides) with all the sequences of a database using the
statistical methods of Karlin and Altschul
Bibliografia:
Strachan T. e Read A.P. Human Molecular Genetics. 3rd edition. Garland Science,
Taylor & Francis Group, 2004
Regateiro F.J. Manual de Gentica Mdica. Imprensa da Universidade, Coimbra,
2003.
Braunwald E, Fauci A, Kasper D, Hauser S, Longo D, Jameson JL. Harrisons
Principles of Internal Medicine. McGraw-Hill 16th Edition. 2005
Lodish H. et al. Molecular Cell Biology. W.H.Freeman and Company. 5
th
edition
Azevedo C. Biologia Celular e Molecular. Lidel. 2005
Genetics Home Reference: http://ghr.nlm.nih.gov/
International HapMap Project, www.hapmap.org
Nobel Prize. http://nobelprize.org/index.html
Applied Biosystems. http://www.ambion.com/techlib/read/
Relatrio n51 do Conselho Nacional de tica para as cincias da Vida,
Presidncia do Conselho de Ministros. disponvel em:
http://www.cnecv.gov.pt/NR/rdonlyres/A81B2738-8A62-458F-90B5-
BEB174EB3A30/0/P051_RelatorioDGPI.pdf
Apresentao de power point sobre a regulao do ciclo celular de Ana Gomes do
Instituto de Anatomia Patolgica da Faculdade de Medicina da Universidade de
Coimbra, 29 Maio de 2009 disponvel em:
https://woc.uc.pt/fmuc/getFile.do?tipo=1&id=319
Lao O, Andrs AM, Mateu E, Bertranpetit J, Calafell F. Spatial patterns of cystic
fibrosis mutation spectra in European populations. European Journal of Human
Genetics (2003) 11, 385394.
Souza MV, Krug BC, PicON PD, Schwartz, IVD. Medicamentos de alto custo para
doenas raras no Brasil: o exemplo das doenas lisossmicas. Revista Cincia &
Sade Coletiva da Associao Brasileira de Ps-Graduao em Sade Coletiva. 2007.
disponvel em:
http://www.abrasco.org.br/cienciaesaudecoletiva/artigos/artigo_int.php?id_artigo=2406
Fairbairn LJ, Lashford LS, Spooncer E, McDermott RH, Lebens G, Arrand JE,
Arrand JR, Bellantuono I, Holt R, Hatton CE, Cooper A, Besley GT, Wraith JE, Anson
DS, Hopwood JJ, Dexter TM. Towards gene therapy of Hurler syndrome. Cas Lek
Cesk. 1997 Jan 8;136(1):27-31.
Anabela Pinto, Madalena Dias, Eugnio Tefilo, Vtor Brotas, M. Ermelinda
Pereira. Variante Btica da Glucose-6-fosfato-desidrogenase: a propsito de um caso
clnico. Medicina Interna, Casos Clnicos. 2005. disponvel em:
http://www.spmi.pt/revista/vol13/vol13_n1_2006_28_32.pdf
Andrea Cristina SantAna, Alessandra Paiva Puertas Alves. Seminrio DNA
mitocondrial. Universidade Federal do Par, Departamento de Patologia. Laboratrio
de Gentica Humana e Mdica- Curso de Especializao em gentica Forense. 2005.
disponvel em: http://www.seguranca.mt.gov.br/politec/3c/artigos/dna_mitocondrial.doc
Mattick JS, Makunin IV. Non-coding RNA. Human Molecular Genetics, 2006, Vol.
15, Review Issue 1
P. P. Van Veldhoven, M. Casteels, G. P. Mannaerts, M. Baes. Further insights into
peroxisomal lipid breakdown via a- and b-oxidation. Biochemical Society Transactions.
2001; 29, (292298)
Nigro JM, Takahashi MA, Ginzinger
DG, Law M, Passe S, Jenkins
RB, K Aldape.
Detection of 1p and 19q Loss in Oligodendroglioma by Quantitative Microsatellite
Analysis, a Real-Time Quantitative Polymerase Chain Reaction Assay. American
Journal of Pathology. 2001;158:1253-1262.)
IB Weinstein, AK Joe. Oncogene Addiction-A Rationale for Molecular Targeting:
Mechanisms of Oncogene Addiction. Nat Clin Pract Oncol. 2006;3(8):448-457 URL:
http://www.medscape.com/viewarticle/543869_4
Stratton MR, Campbell PJ, & Futreal PA. The cancer genome. Nature 458, 719-
724(9 April 2009). URL:
http://www.nature.com/nature/journal/v458/n7239/box/nature07943_BX1.html
McNeil N, Montagna C, Difilippantonio MJ and Ried T . Comparative Cancer
Cytogenetics. Atlas Genet Cytogenet Oncol Haematol. August 2003.
URL : http://AtlasGeneticsOncology.org/Deep/ComparCancerCytogID20011.html
http://medicosdeportugal.saude.sapo.pt/action/0/
http://emedicine.medscape.com/article/983678-overview
http://www.gistsupport.org/media/cells%20%20DNA/cellcycle_CTI_575pix.jpg
You might also like
- Aula - Genética Do Câncer PDFDocument70 pagesAula - Genética Do Câncer PDFAna OlindaNo ratings yet
- Aula 3 - Cards - BASES MOLECULARES DEL CÁNCERDocument5 pagesAula 3 - Cards - BASES MOLECULARES DEL CÁNCERruan loiolaNo ratings yet
- Erica SSAraujoDocument210 pagesErica SSAraujoJoão LukasNo ratings yet
- Genetica 1 FrequenciaDocument29 pagesGenetica 1 FrequenciaAndréia SilvaNo ratings yet
- Lacetilcarnitina en Tratamiento Hiperactividad X FragilDocument6 pagesLacetilcarnitina en Tratamiento Hiperactividad X Fragilmariniuxrd1467No ratings yet
- Genetica SebentaDocument102 pagesGenetica SebentaInês Ferreira100% (1)
- Revisão Ted 2011 - Casos Relevantes para EstudoDocument142 pagesRevisão Ted 2011 - Casos Relevantes para EstudoAndré Cartell100% (1)
- Patologia - Resumo Robbins 5 - Doenças GenéticasDocument12 pagesPatologia - Resumo Robbins 5 - Doenças Genéticasrcnrsp3850No ratings yet
- Revisão Literária STDocument3 pagesRevisão Literária STluisaisabella399No ratings yet
- RESUMO DOS EDs - GENÉTICA PROVA 2Document8 pagesRESUMO DOS EDs - GENÉTICA PROVA 2Larissa Sayuri YoshizawaNo ratings yet
- Aps Genética - Final PDFDocument12 pagesAps Genética - Final PDFjlribeiro1315No ratings yet
- Resumo Doencas GeneticasDocument12 pagesResumo Doencas Geneticasdanilo186No ratings yet
- Lista 9 RespondidaDocument5 pagesLista 9 RespondidaAna Crystina MaiaNo ratings yet
- Livro Texto - Unidade IIIDocument82 pagesLivro Texto - Unidade IIIosakatacorrea030No ratings yet
- PPG (Roteiro - Aula 6) - Bases Moleculares Do CâncerDocument4 pagesPPG (Roteiro - Aula 6) - Bases Moleculares Do Câncertuts1704No ratings yet
- Hereditariedade MitocondrialDocument7 pagesHereditariedade MitocondrialEliana RodriguesNo ratings yet
- 16 - Epigenética e ComportamentoDocument23 pages16 - Epigenética e ComportamentojosemscastroNo ratings yet
- 39 Leucemia Mieloide Cronica Diagnostico e TratamentoDocument30 pages39 Leucemia Mieloide Cronica Diagnostico e TratamentoAdriana ConstantNo ratings yet
- Relatório - Ase 2.problema 04.tutoria BDocument7 pagesRelatório - Ase 2.problema 04.tutoria BFernanda HerênioNo ratings yet
- Aula 1 - Processos PatológicosDocument34 pagesAula 1 - Processos PatológicosGiovanna Juliani SofiattiNo ratings yet
- Estágio de Citogenética2Document56 pagesEstágio de Citogenética2Fenias BoaneNo ratings yet
- PolimorfismoDocument83 pagesPolimorfismoGabileteNo ratings yet
- Capítulo 6 Robbins PatologiaDocument10 pagesCapítulo 6 Robbins PatologiaCarolina StrongerNo ratings yet
- Genetica MedicaDocument3 pagesGenetica Medicapedro.valves03No ratings yet
- Resumo DCCDocument61 pagesResumo DCCJohnny MendesNo ratings yet
- CCRDocument23 pagesCCRJulio César Campos LópezNo ratings yet
- Livro de Genética - Unidade 4 (Zigotos435)Document24 pagesLivro de Genética - Unidade 4 (Zigotos435)tenarthur01No ratings yet
- Artigo - Linf Cel T - NK - VSTCCDocument8 pagesArtigo - Linf Cel T - NK - VSTCCMarina OliveiraNo ratings yet
- Base Molecular Do Cancer Parte 1Document7 pagesBase Molecular Do Cancer Parte 1ÁgathaNo ratings yet
- Nova Geração DissertaçãoDocument41 pagesNova Geração Dissertaçãorafaela SalesNo ratings yet
- Estudo Dirido de Herpes e Papilomavirus Humanos. João Pedro Monteiro.Document5 pagesEstudo Dirido de Herpes e Papilomavirus Humanos. João Pedro Monteiro.Joao Pedro MonteiroNo ratings yet
- PoliposeDocument114 pagesPoliposesraimundo_1No ratings yet
- Projeto PUB IB 2022-2023Document6 pagesProjeto PUB IB 2022-2023ceu ceuNo ratings yet
- Gen 2Document14 pagesGen 2boraquicarNo ratings yet
- Sequenciação de GenomasDocument6 pagesSequenciação de GenomasMariana NetoNo ratings yet
- Aula 1Document13 pagesAula 1Sérgio FonsecaNo ratings yet
- Aula 3.1Document36 pagesAula 3.1beatrizgabriellenunesNo ratings yet
- Bio 2024Document2 pagesBio 2024claudio.filhoNo ratings yet
- CARCINOGÊNESE - ResumoDocument6 pagesCARCINOGÊNESE - ResumoJulyanne GoesNo ratings yet
- Aula Genética Do CancerDocument36 pagesAula Genética Do CancerJullyara UlhNo ratings yet
- Caso 3Document13 pagesCaso 3anna.vieiraNo ratings yet
- Leucemias Agudas (Capítulo de Livro)Document27 pagesLeucemias Agudas (Capítulo de Livro)lovatozueraNo ratings yet
- Alteracoes Do Material Genetico PDFDocument68 pagesAlteracoes Do Material Genetico PDFRicardo Batista100% (1)
- Conferência - 10 - Mieloma MúltiploDocument3 pagesConferência - 10 - Mieloma MúltiploMarcos Fernandes MendesNo ratings yet
- Genética e EmbriologiaDocument12 pagesGenética e EmbriologiadadizachiNo ratings yet
- Bases Geneticas Das DoencasDocument17 pagesBases Geneticas Das DoencasAndreutt TabosaNo ratings yet
- Provas Teóricas ProliferaçãoDocument81 pagesProvas Teóricas Proliferaçãolaissa.daveigalustosaNo ratings yet
- Curso Ouro Preto-Serie Branca UltimoDocument166 pagesCurso Ouro Preto-Serie Branca Ultimoray_limaaNo ratings yet
- Relatório BioinformáticaDocument15 pagesRelatório BioinformáticaÍris AlvesNo ratings yet
- Linfossarcoma e Sindrome de Ma Absorcao - Michele de Menezes TruppelDocument41 pagesLinfossarcoma e Sindrome de Ma Absorcao - Michele de Menezes TruppelJORGE_46693822No ratings yet
- Genética Humana (Completa!!!)Document186 pagesGenética Humana (Completa!!!)Viviane Medeiros Silveira100% (2)
- Famed Plano de Ensino McgnoDocument5 pagesFamed Plano de Ensino McgnoAriadna ZambelliNo ratings yet
- Carcinogênese 2023Document57 pagesCarcinogênese 2023Ana Livia CamolesiNo ratings yet
- Artigo Oncologia RevisãoDocument8 pagesArtigo Oncologia RevisãoAlessia NunesNo ratings yet
- Leucemia Mielóide Crônica e o Sistema Fas-FasLDocument6 pagesLeucemia Mielóide Crônica e o Sistema Fas-FasLRenata AmaralNo ratings yet
- Teste de Progresso 2016 - ComentadoDocument20 pagesTeste de Progresso 2016 - ComentadoLeandroFrancoNo ratings yet
- Assuntos ObbDocument5 pagesAssuntos ObbCapuama AndersonNo ratings yet
- Edital Verticalizado - Henrique Lobel - PDF - 231226 - 173343Document33 pagesEdital Verticalizado - Henrique Lobel - PDF - 231226 - 173343julia.starling.neg1No ratings yet
- 284-2028-1-PB ResinasDocument6 pages284-2028-1-PB ResinasMiguelzip2000No ratings yet
- Características Fisicoquímicas e Citológicas Do Líquido Sinovial Da Articulação Temporomandibular em EqüinosDocument5 pagesCaracterísticas Fisicoquímicas e Citológicas Do Líquido Sinovial Da Articulação Temporomandibular em EqüinosMárcia LoureiroNo ratings yet
- Introdução A Cirurgia - CópiaDocument73 pagesIntrodução A Cirurgia - CópiaLucas Silva100% (1)
- Feng ShuiDocument9 pagesFeng ShuiMiguelzip2000No ratings yet
- Características Fisicoquímicas e Citológicas Do Líquido Sinovial Da Articulação Temporomandibular em EqüinosDocument5 pagesCaracterísticas Fisicoquímicas e Citológicas Do Líquido Sinovial Da Articulação Temporomandibular em EqüinosMárcia LoureiroNo ratings yet
- JBC 25Document11 pagesJBC 25Claudio BrittoNo ratings yet
- En Do Don Ti ADocument35 pagesEn Do Don Ti AKyze QuintelaNo ratings yet
- Introdução A Neurologia VeterináriaDocument37 pagesIntrodução A Neurologia VeterináriaIgor Saldanha de FreitasNo ratings yet
- Sebenta Anestesia Volátil e MonitorizaçãoDocument50 pagesSebenta Anestesia Volátil e MonitorizaçãoMiguelzip2000No ratings yet
- Exercícios Genealogia 2Document1 pageExercícios Genealogia 2Miguelzip2000No ratings yet
- Exercícios Genealogia 1Document1 pageExercícios Genealogia 1Miguelzip2000No ratings yet
- AnestesiologiaDocument83 pagesAnestesiologiaMiguelzip2000No ratings yet
- Exercícios de Genealogia e TransmissãoDocument2 pagesExercícios de Genealogia e TransmissãoMiguelzip2000No ratings yet
- ImpressõesDocument4 pagesImpressõesMiguelzip2000No ratings yet
- BodyDocument29 pagesBodyMiguelzip2000No ratings yet
- Exercícios Transmissão e Calculo RiscoDocument6 pagesExercícios Transmissão e Calculo RiscoMiguelzip2000No ratings yet
- SBV Sebenta MinhaDocument27 pagesSBV Sebenta MinhaMiguelzip2000No ratings yet
- Circulação de Animais CompanhiaDocument15 pagesCirculação de Animais CompanhiaMiguelzip2000No ratings yet
- Anestesia 1Document31 pagesAnestesia 1Miguelzip2000No ratings yet
- Materia para Oral de MorfologiaDocument1 pageMateria para Oral de MorfologiaMiguelzip2000No ratings yet
- 1882 - Almanak Do Ministerio Da GuerraDocument389 pages1882 - Almanak Do Ministerio Da GuerraSamuel Robes LoureiroNo ratings yet
- 4 BI A Braudel Repensando A Vida MaterialDocument69 pages4 BI A Braudel Repensando A Vida MaterialDalton AndradeNo ratings yet
- Manual de Segurança e Boas Práticas INEMDocument224 pagesManual de Segurança e Boas Práticas INEMRúben VianaNo ratings yet
- A Metodologia Design Thinking Como Estratégia Gerencial para Empreendimentos (Revisar)Document19 pagesA Metodologia Design Thinking Como Estratégia Gerencial para Empreendimentos (Revisar)Wanderson SantosNo ratings yet
- Miasmas Luiz PDFDocument37 pagesMiasmas Luiz PDFLuiz Arias100% (1)
- CORCORAN, N. Teorias e Modelos Na Comunicação de Mensagens de SaúdeDocument15 pagesCORCORAN, N. Teorias e Modelos Na Comunicação de Mensagens de SaúdeÁdria AlbaradoNo ratings yet
- Tecumseh Palestra 2015Document131 pagesTecumseh Palestra 2015Marcus Banzatto100% (1)
- O Dom de ProfeciaDocument116 pagesO Dom de ProfeciaLeo IgildoNo ratings yet
- Livro Texto - Unidade IIDocument61 pagesLivro Texto - Unidade IIGustavo NascimentoNo ratings yet
- Prova Integrado Picos Salas 4 e 5 2019-12-01Document12 pagesProva Integrado Picos Salas 4 e 5 2019-12-01Fabio AlvesNo ratings yet
- Razão Vontade e DesejoDocument12 pagesRazão Vontade e DesejoEduardo DumkeNo ratings yet
- Outubro Rosa CA de MamaDocument11 pagesOutubro Rosa CA de MamaCPSST100% (1)
- Edital ValinhosDocument80 pagesEdital Valinhosdiogomelop2193No ratings yet
- 6 Tipagem ABO - 13set2016Document27 pages6 Tipagem ABO - 13set2016Adriana Silva100% (1)
- Unidade de Medidas - Fisica 1 AnoDocument4 pagesUnidade de Medidas - Fisica 1 AnoClaudemir NunesNo ratings yet
- Teste Circuito Económico 1Document7 pagesTeste Circuito Económico 1Sara Ferrão100% (1)
- Demonstracões Financeiras EZTEC 2019Document50 pagesDemonstracões Financeiras EZTEC 2019Diego PiresNo ratings yet
- Mapa Mental CriminologiaDocument11 pagesMapa Mental CriminologiaJuliana Cordeiro0% (1)
- Estudo de Caso - Hospital SamaritanoDocument2 pagesEstudo de Caso - Hospital SamaritanoNelma Maria AlvesNo ratings yet
- Brma Install ManualDocument58 pagesBrma Install ManualLucasAndreNo ratings yet
- Resumo HistoplasmoseDocument5 pagesResumo HistoplasmoseGustavo I. Moraes100% (1)
- A6 - Circuito RCDocument7 pagesA6 - Circuito RCVinicius MoraesNo ratings yet
- Re 82176 Hca10 Aula 16Document8 pagesRe 82176 Hca10 Aula 16Bruno HenriquesNo ratings yet
- HTTP Culto Jovem Opçoes de JaDocument7 pagesHTTP Culto Jovem Opçoes de JajorgenacNo ratings yet
- Cinesioterapia (Aula 4 - Postura)Document51 pagesCinesioterapia (Aula 4 - Postura)ketsume.wsNo ratings yet
- Emprego Das Classes de Palavras 1Document10 pagesEmprego Das Classes de Palavras 1Rodolfo SilvaNo ratings yet
- Empreendedorismo Avaliação On-Line 1 (AOL 1) - QuestionárioDocument13 pagesEmpreendedorismo Avaliação On-Line 1 (AOL 1) - QuestionárioMarina Aguiar MoraisNo ratings yet
- Curso 1Document6 pagesCurso 1JR JRNo ratings yet
- DCR Aimorés 2017Document33 pagesDCR Aimorés 2017Lusineide SilvaNo ratings yet
- Iniciaçao Reiki 3BDocument8 pagesIniciaçao Reiki 3BMarioMoraisNo ratings yet