You might also like
- Seduction Theory - The NameDocument20 pagesSeduction Theory - The Nameharrridan100% (1)
- Group Therapy Techniques Derived from PsychoanalysisDocument12 pagesGroup Therapy Techniques Derived from PsychoanalysisJuanMejiaNo ratings yet
- Excerpted From "Hallucinations" by Oliver Sacks. Copyright © 2013 by Oliver Sacks. With Permission of The Publisher, Vintage.Document31 pagesExcerpted From "Hallucinations" by Oliver Sacks. Copyright © 2013 by Oliver Sacks. With Permission of The Publisher, Vintage.wamu885No ratings yet
- The Freudian CutDocument16 pagesThe Freudian CutRichard G. KleinNo ratings yet
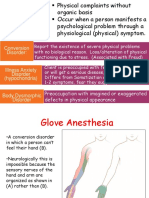
- Somatic Symptom Disorders and Personality DisordersDocument15 pagesSomatic Symptom Disorders and Personality Disordersapi-287575009No ratings yet
- GibsonTheory of Percetual LearningDocument10 pagesGibsonTheory of Percetual LearningAlfonso García VelázquezNo ratings yet
- The End of Narcissism: Re-Visioning The Oceanic FeelingDocument11 pagesThe End of Narcissism: Re-Visioning The Oceanic FeelingChristopher CordryNo ratings yet
- Cognition, Brain, and Consciousness: Introduction to Cognitive NeuroscienceFrom EverandCognition, Brain, and Consciousness: Introduction to Cognitive NeuroscienceRating: 4 out of 5 stars4/5 (4)
- Shai-Fonagy Parental Embodied MentalizingDocument26 pagesShai-Fonagy Parental Embodied MentalizingCorina IoanaNo ratings yet
- Melanie Klein Origins of TransferanceDocument8 pagesMelanie Klein Origins of TransferanceVITOR HUGO LIMA BARRETONo ratings yet
- Finding The Self in MindDocument15 pagesFinding The Self in MindAlbert Muya MurayaNo ratings yet
- The Fiction of Memory-Elizabeth LoftusDocument4 pagesThe Fiction of Memory-Elizabeth LoftusUma MaheswariNo ratings yet
- Fonagy 1999Document16 pagesFonagy 1999sfdkhkfjsfnsk njfsnksfnNo ratings yet
- Izard (2009) Emotion TheoryDocument29 pagesIzard (2009) Emotion Theoryred_cocoa96No ratings yet
- 1951 Searles Manifestations of IncorporationDocument31 pages1951 Searles Manifestations of IncorporationKevin McInnesNo ratings yet
- Reflections On ShameDocument21 pagesReflections On ShameAnamaria TudoracheNo ratings yet
- Scharff Sobre FairbairnDocument21 pagesScharff Sobre FairbairnAntonio TariNo ratings yet
- Memory DysfunctionDocument8 pagesMemory DysfunctionAdamarloNo ratings yet
- On Script Theory and Disgust PsychologyDocument29 pagesOn Script Theory and Disgust PsychologySid SudiacalNo ratings yet
- Discovering Meaning in PanicDocument445 pagesDiscovering Meaning in Paniclotaoto100% (1)
- A Neuropsychoanalytic Viewpoint: Commentary On Paper by Steven H. KnoblauchDocument26 pagesA Neuropsychoanalytic Viewpoint: Commentary On Paper by Steven H. KnoblauchPaul ScallanNo ratings yet
- FreudDocument2 pagesFreudLester AguirreNo ratings yet
- "Finding My Own Identity" - A Qualitative Metasynthesis of Adult Anorexia Nervosa Treatment ExperiencesDocument14 pages"Finding My Own Identity" - A Qualitative Metasynthesis of Adult Anorexia Nervosa Treatment ExperiencesAnastasia DoyenNo ratings yet
- Neural Basis of ConsciousnessDocument13 pagesNeural Basis of ConsciousnessAlejandro León100% (1)
- Winnicott ch1 PDFDocument18 pagesWinnicott ch1 PDFAlsabila NcisNo ratings yet
- Special Article: Biology and The Future of Psychoanalysis: A New Intellectual Framework For Psychiatry RevisitedDocument20 pagesSpecial Article: Biology and The Future of Psychoanalysis: A New Intellectual Framework For Psychiatry RevisitedFelipeHenriquezRuz100% (1)
- Bornstein, B. - On LatencyDocument8 pagesBornstein, B. - On LatencycabaretdadaNo ratings yet
- Clewell, Shades of Mourning in Kieslowski's Three Colours TrilogyDocument8 pagesClewell, Shades of Mourning in Kieslowski's Three Colours TrilogydiapappNo ratings yet
- Tustin (1994) - Perpetuation of An Error. Journal of Child Psychotherapy.20 (1), Pp.3-23Document22 pagesTustin (1994) - Perpetuation of An Error. Journal of Child Psychotherapy.20 (1), Pp.3-23Lucia SalasNo ratings yet
- Masochism+A +connolyDocument21 pagesMasochism+A +connolyAnonymous KgUtPlkjNo ratings yet
- Creation of The Self and Language - Primitive Sensory RelationsDocument80 pagesCreation of The Self and Language - Primitive Sensory RelationsSofi LlorensNo ratings yet
- Not To Forget Tom Andersen Way of BeingDocument20 pagesNot To Forget Tom Andersen Way of BeingignNo ratings yet
- The dead mother syndrome and its effects on sexual developmentDocument1 pageThe dead mother syndrome and its effects on sexual developmentCiprian Cătălin AlexandruNo ratings yet
- Maken Sense - HinshelwoodDocument13 pagesMaken Sense - HinshelwoodEver KarNo ratings yet
- Louvet Creators and Szondi Test PDFDocument299 pagesLouvet Creators and Szondi Test PDFVicentiu MafteiNo ratings yet
- Fdocuments - in - The British Object Relations Winnicott and Fairbairn Chapter 5 PDFDocument15 pagesFdocuments - in - The British Object Relations Winnicott and Fairbairn Chapter 5 PDFLu GalvisNo ratings yet
- Damasio Emotions and Decision MakingDocument13 pagesDamasio Emotions and Decision MakingwisjanuszNo ratings yet
- Anger and Stress ManagementDocument40 pagesAnger and Stress ManagementMaharaja MaharongaNo ratings yet
- Craig InsulaDocument12 pagesCraig InsulaJúlia PriyaNo ratings yet
- Unconscious Facial Reactions To Emotional Facial ExpressionsDocument5 pagesUnconscious Facial Reactions To Emotional Facial ExpressionsLigia AndreiNo ratings yet
- Liotti Trauma Attachment 2004Document39 pagesLiotti Trauma Attachment 2004samathacalmmindNo ratings yet
- PEP Web - Constructions in AnalysisDocument7 pagesPEP Web - Constructions in Analysisdsh265No ratings yet
- The Mind and The Brain: Neuroplasticity and The Power of Mental Force - Jeffrey M. Schwartz MDDocument5 pagesThe Mind and The Brain: Neuroplasticity and The Power of Mental Force - Jeffrey M. Schwartz MDwybupody0% (1)
- Pete Walker, M.A. PsychotherapyDocument1 pagePete Walker, M.A. PsychotherapyDušana NikolićNo ratings yet
- Susanna Isaacs ElmhirstDocument14 pagesSusanna Isaacs Elmhirstkarine botelhoNo ratings yet
- Somatic Transference and Countertransference in Psychoanalytic Intersubjective Dance/Movement TherapyDocument17 pagesSomatic Transference and Countertransference in Psychoanalytic Intersubjective Dance/Movement TherapyMaría Elisa Al Cheikh StrubingerNo ratings yet
- Attachment and Loss Retrospect and Prospect BowblyDocument15 pagesAttachment and Loss Retrospect and Prospect BowblyVirginia SummerNo ratings yet
- Falling in Love with Love: Oedipal and Post Oedipal ManifestationsDocument28 pagesFalling in Love with Love: Oedipal and Post Oedipal ManifestationsIrit PazNo ratings yet
- Oxford Handbooks Online: A Better Self: Freud and Nietzsche On The Nature and Value of SublimationDocument978 pagesOxford Handbooks Online: A Better Self: Freud and Nietzsche On The Nature and Value of SublimationJose BusaidNo ratings yet
- Internal Racism A Psychoanalytic Approach To Race and Difference, Stephens & Alanson 2014Document3 pagesInternal Racism A Psychoanalytic Approach To Race and Difference, Stephens & Alanson 2014Ambrose66100% (1)
- Hypnotic Intervention For Ambiguity As A Depressive Risk FactorDocument9 pagesHypnotic Intervention For Ambiguity As A Depressive Risk FactorOespiritodedeus MestreNo ratings yet
- Peter Fonagy The Development of Psychopathology From Infancy To AdulthoodDocument29 pagesPeter Fonagy The Development of Psychopathology From Infancy To AdulthoodjuaromerNo ratings yet
- 3 - Transforming Normality Into Pathology PDFDocument13 pages3 - Transforming Normality Into Pathology PDFAnonymous quHwGv1jvnNo ratings yet
- Melancholia in a Sixteenth-Century Spanish NunDocument5 pagesMelancholia in a Sixteenth-Century Spanish NunatabeiNo ratings yet
- Memory Diaspora and HysteriaDocument27 pagesMemory Diaspora and HysteriamgterNo ratings yet
- D.LAİNG Mystification, Confusion and ConflictDocument1 pageD.LAİNG Mystification, Confusion and Conflictneo-kerataNo ratings yet
- On Psychoanalysis of Autism: How Our Mind Works In Social ImpairmentFrom EverandOn Psychoanalysis of Autism: How Our Mind Works In Social ImpairmentNo ratings yet
- Amnesia: Clinical, Psychological and Medicolegal AspectsFrom EverandAmnesia: Clinical, Psychological and Medicolegal AspectsC. W. M. WhittyNo ratings yet
- On Literature, Revolution, Entropy and Other Matters - Yevegney ZamyatinDocument4 pagesOn Literature, Revolution, Entropy and Other Matters - Yevegney ZamyatinRachel AHNo ratings yet
- Capitalism's Traumatic Encounter With LackDocument16 pagesCapitalism's Traumatic Encounter With LackRachel AHNo ratings yet
- Reveries of The Solitary Walker, RousseauDocument11 pagesReveries of The Solitary Walker, RousseauRachel AHNo ratings yet
- Zen Poems, Read by Lucien StrykDocument10 pagesZen Poems, Read by Lucien StrykRachel AHNo ratings yet
- Philosophy and The Black PanthersDocument7 pagesPhilosophy and The Black PanthersRachel AH100% (1)
- Revolutionary SuicideDocument182 pagesRevolutionary SuicideRachel AH100% (3)
- Battcock The Art of Performance 1984Document152 pagesBattcock The Art of Performance 1984carrapatoso100% (1)
- Pussy Riot Closing StatementsDocument11 pagesPussy Riot Closing StatementsRachel AHNo ratings yet
- Melancholy and The Care of The Soul IntroDocument17 pagesMelancholy and The Care of The Soul IntroIvanKovačNo ratings yet
- Total FearsDocument21 pagesTotal FearsRachel AH100% (1)
- The Love Suicides at AmijimaDocument32 pagesThe Love Suicides at AmijimaRachel AHNo ratings yet
- The Tragedy of Schizophrenia Without PsychotherapyDocument30 pagesThe Tragedy of Schizophrenia Without PsychotherapyRachel AHNo ratings yet
- The Ominous NuminousDocument18 pagesThe Ominous NuminousRachel AH0% (1)
- The Secret History of The MongolsDocument172 pagesThe Secret History of The MongolsRachel AH100% (5)
- Notes From The Underground TDocument203 pagesNotes From The Underground TCTpatelNo ratings yet
- Tales of The Floating WorldDocument163 pagesTales of The Floating WorldRachel AHNo ratings yet