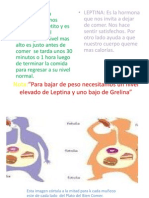
You might also like
- Gordos y FlacosDocument27 pagesGordos y FlacosTati Del MarNo ratings yet
- Marco Teorico 1Document29 pagesMarco Teorico 1Mario Perez RumboNo ratings yet
- Consumo voluntario de nutrimentos en animales domésticosDocument6 pagesConsumo voluntario de nutrimentos en animales domésticosERIK AXEL CRUZ ALCANTARNo ratings yet
- 1er Articulo-Nutrigenomica y ObesidadDocument4 pages1er Articulo-Nutrigenomica y ObesidadJOSE ISAAC SANTOS CONTRERASNo ratings yet
- Consumo VoluntarioDocument8 pagesConsumo VoluntarioPatrick Fernando Weill VilcaNo ratings yet
- Consumo VoluntarioDocument27 pagesConsumo VoluntarioMarielena GarciaNo ratings yet
- 327729-Texto Del Art - Culo-155021-5-10-20210426Document11 pages327729-Texto Del Art - Culo-155021-5-10-20210426Vėrónïcå CuėncåNo ratings yet
- Consumo VoluntarioDocument10 pagesConsumo VoluntarioonenehoNo ratings yet
- 3b Control Del Hambre y ApetitoDocument36 pages3b Control Del Hambre y ApetitoKarla MirellNo ratings yet
- Hambre y Saciedad - En.esDocument7 pagesHambre y Saciedad - En.esMelanyNo ratings yet
- Centros Nerviosos de Hambre y SaciedadDocument39 pagesCentros Nerviosos de Hambre y SaciedadrolandoNo ratings yet
- Clase 2 REGULACION DE LA CONDUCTA ALIMENTARIADocument32 pagesClase 2 REGULACION DE LA CONDUCTA ALIMENTARIARenato Ibañez MusayónNo ratings yet
- Grelina ResumenDocument3 pagesGrelina ResumenMaría Laura ZarazúaNo ratings yet
- REGULADORESNEUROENDOCRINOSYGASTROINTESTINALESDELAPETITOYLASACIEDADDocument28 pagesREGULADORESNEUROENDOCRINOSYGASTROINTESTINALESDELAPETITOYLASACIEDADBarbaraNo ratings yet
- Regulacion Hormonal Gastrointestinal Pequeños RumiantesDocument9 pagesRegulacion Hormonal Gastrointestinal Pequeños RumiantesDana SandovalNo ratings yet
- Guia Examen 1°Document46 pagesGuia Examen 1°eunice09veraNo ratings yet
- RP7 VaciamientoDocument5 pagesRP7 VaciamientoraymundoNo ratings yet
- Articulo Sobre Fase Cefalica en La DigestionDocument15 pagesArticulo Sobre Fase Cefalica en La DigestionJaquelin PeraltaNo ratings yet
- Monografia Nutrigenómica y ObesidadDocument34 pagesMonografia Nutrigenómica y ObesidadRenzo ReyanNo ratings yet
- Fisiologia de La SaciedadDocument5 pagesFisiologia de La SaciedadFran Villar HormazábalNo ratings yet
- Mecanismos Regulatorios de La IngestiónDocument6 pagesMecanismos Regulatorios de La IngestiónLihoy KamNo ratings yet
- Hormonas GastronitesDocument14 pagesHormonas GastronitesAbril VelásquezNo ratings yet
- Ayudantía 05Document17 pagesAyudantía 05Claudia PizarroNo ratings yet
- Hambre, Sed y SaciedadDocument22 pagesHambre, Sed y SaciedadCitlalli OsirisNo ratings yet
- Metabolismo en El AyunoDocument10 pagesMetabolismo en El AyunoMaricris_85No ratings yet
- HAMBRE ApuntesDocument10 pagesHAMBRE ApuntesSoraya ShurairNo ratings yet
- Anorexia y Bulimia Nerviosa - ResumenDocument3 pagesAnorexia y Bulimia Nerviosa - ResumenEmiliano ParedesNo ratings yet
- 2003 n1 Revision Regulacion Del Apetito Nuevos ConceptosDocument8 pages2003 n1 Revision Regulacion Del Apetito Nuevos ConceptosFederico TedescoNo ratings yet
- 3 Emprend Ayuno 1631666879680Document27 pages3 Emprend Ayuno 1631666879680Jany PiccoNo ratings yet
- Fisiologia Hormonal IntestinalDocument14 pagesFisiologia Hormonal IntestinalChristian Benjo Lopez EidNo ratings yet
- Nutrición Deportiva PDFDocument121 pagesNutrición Deportiva PDFJonatan HernandézNo ratings yet
- Fisiologia Uabp7Document5 pagesFisiologia Uabp7Santiago SimonielloNo ratings yet
- Npnu U1 A2 DiraDocument5 pagesNpnu U1 A2 DiradianaNo ratings yet
- M2-T4 Regulación Fisiológica de La AlimentaciónDocument10 pagesM2-T4 Regulación Fisiológica de La AlimentaciónSil SerranoNo ratings yet
- Estómagos - ¿Importa El Tamaño - Aspectos de Saciedad Intestinal, Saciedad Gástrica, Hambre y GulaDocument4 pagesEstómagos - ¿Importa El Tamaño - Aspectos de Saciedad Intestinal, Saciedad Gástrica, Hambre y GulaNutrition NationNo ratings yet
- Alfredo Hormonas RegulacionDocument65 pagesAlfredo Hormonas RegulacionMardabNo ratings yet
- Grelina vs Leptina: las hormonas del hambre y la saciedadDocument7 pagesGrelina vs Leptina: las hormonas del hambre y la saciedadMelina Pacheco MendiolaNo ratings yet
- Monografía Motivación y EmociónDocument12 pagesMonografía Motivación y EmociónGary KruzNo ratings yet
- Fisiologia Hambre ApetitoDocument60 pagesFisiologia Hambre ApetitoArtur CrossNo ratings yet
- Ciclo Alimento AyunoDocument9 pagesCiclo Alimento AyunoFredy JrNo ratings yet
- Factores que modifican la conducta alimentariaDocument3 pagesFactores que modifican la conducta alimentariaCarolina Arroyo Yépez.No ratings yet
- RESUMENDocument2 pagesRESUMENjuanmideluxeNo ratings yet
- Teoría GlucostaticaDocument17 pagesTeoría GlucostaticaMichelleNo ratings yet
- Biopsicología 12 HambreDocument5 pagesBiopsicología 12 HambreEmmanuel Domínguez RosalesNo ratings yet
- Guía 21 RespuestasDocument3 pagesGuía 21 Respuestasramirezmisa2612No ratings yet
- Vaciamiento GastricoDocument4 pagesVaciamiento GastricoarelyNo ratings yet
- ObecidadDocument26 pagesObecidadMao SolanoNo ratings yet
- Cirugía Bariátrica Qué Papel Juegan Las Hormonas GastrointestinalesDocument12 pagesCirugía Bariátrica Qué Papel Juegan Las Hormonas GastrointestinalesefallxNo ratings yet
- Alimentos Funcionales y ObesidadDocument19 pagesAlimentos Funcionales y Obesidadtheseal857No ratings yet
- 09 EDV475735.inddDocument15 pages09 EDV475735.inddYarely De la RosaNo ratings yet
- Cuestionario de ComerDocument3 pagesCuestionario de ComerIdelsa KellyNo ratings yet
- Conductas IngestivasDocument54 pagesConductas IngestivasFernanda PérezNo ratings yet
- Entradas Y Salidas Energeticas Equilibradas en Condiciones EstacionariasDocument10 pagesEntradas Y Salidas Energeticas Equilibradas en Condiciones EstacionariasDIEGO CRISTHOPER MALPARTIDA ESPINOZANo ratings yet
- Juliana Heno-Tarea EMBARAZO Y METABOLISMODocument15 pagesJuliana Heno-Tarea EMBARAZO Y METABOLISMOJuliana HenaoNo ratings yet
- Alimentos Funcionales Que Causan Saciedad 4Document9 pagesAlimentos Funcionales Que Causan Saciedad 4proteinayumNo ratings yet
- 5 Nutricion Enteral + InvestigaciónDocument19 pages5 Nutricion Enteral + InvestigaciónHarold Brayan Ferrufino SejasNo ratings yet
- Vaciamiento GástricoDocument7 pagesVaciamiento GástricoAxelGomezRgNo ratings yet
- Dieta Cetogénica : Dieta Cetogénica Deliciosa Y Fácil Para Una Vida Sana Para Perder PesoFrom EverandDieta Cetogénica : Dieta Cetogénica Deliciosa Y Fácil Para Una Vida Sana Para Perder PesoNo ratings yet
- Alimentación Saludable: La Guía Definitiva: Colección Vida Equilibrada, #4From EverandAlimentación Saludable: La Guía Definitiva: Colección Vida Equilibrada, #4No ratings yet
- Desordenes Del Ciclo Menstrual2016Document43 pagesDesordenes Del Ciclo Menstrual2016Ronald Taboada ZamoraNo ratings yet
- 5-15 Respiracion y Circulacion FetalDocument11 pages5-15 Respiracion y Circulacion FetalJohn Jefferson Angulo CastroNo ratings yet
- Adenopatia RetroauricularDocument6 pagesAdenopatia RetroauricularRonald Taboada ZamoraNo ratings yet
- Endocrino III ObesidadDocument10 pagesEndocrino III ObesidadRonald Taboada ZamoraNo ratings yet
- PalouDocument13 pagesPalouIVANOVICH357No ratings yet
- Endocrino III DiabetesDocument21 pagesEndocrino III DiabetesRonald Taboada ZamoraNo ratings yet
- Informe Succinato DeshidrogenasaDocument7 pagesInforme Succinato DeshidrogenasaStiven SevillaNo ratings yet
- 2inh Incrustación RE5789SCW PDFDocument6 pages2inh Incrustación RE5789SCW PDFluis rivasNo ratings yet
- Caso Proyecto Empresa de Arriendo de Vehículos GDP1463 (1) - TaggedDocument7 pagesCaso Proyecto Empresa de Arriendo de Vehículos GDP1463 (1) - TaggedGONZALO JACOB PROVOSTE MASIASNo ratings yet
- Eda3 1ero Ccss 2023 Prof. Janet OrtegaDocument9 pagesEda3 1ero Ccss 2023 Prof. Janet OrtegaHenry JacobiNo ratings yet
- Dotación agua 38Document18 pagesDotación agua 38denisNo ratings yet
- Aceites y Grasa Insaturados en La Industria de LaDocument4 pagesAceites y Grasa Insaturados en La Industria de LaStefani Ucsa SantoyoNo ratings yet
- El huevo de obsidiana: efectos energéticos y terapéuticosDocument10 pagesEl huevo de obsidiana: efectos energéticos y terapéuticosLupita CuevasNo ratings yet
- Principales Figuras GeometricasDocument5 pagesPrincipales Figuras GeometricasJoche ZamoraNo ratings yet
- Hercampuri VademecumDocument7 pagesHercampuri VademecumJarek5093100% (1)
- Movimientos de Rebaños Ovinos y CaprinosDocument10 pagesMovimientos de Rebaños Ovinos y CaprinosedgarherrerapheNo ratings yet
- Procesos TecnológicosDocument3 pagesProcesos TecnológicosanatellezNo ratings yet
- Plan de Trabajo - Unidad 2 - Bme PDFDocument17 pagesPlan de Trabajo - Unidad 2 - Bme PDFClaudia Osorio FalconiNo ratings yet
- Ampliación y Mejoramiento de La Oferta de Servicios Educativos en 206 Rayito de SolDocument115 pagesAmpliación y Mejoramiento de La Oferta de Servicios Educativos en 206 Rayito de SolCerres Windel100% (1)
- Una Comunidad Es Un Grupo o Conjunto de IndividuosDocument5 pagesUna Comunidad Es Un Grupo o Conjunto de Individuosdelvis_reyes854702100% (1)
- We Bare BearsDocument9 pagesWe Bare BearsYasmin ViverosNo ratings yet
- Curso de Epistemología y Metodología de Investigación CuantitativaDocument78 pagesCurso de Epistemología y Metodología de Investigación Cuantitativadantefabian13No ratings yet
- Planificación de actividades para el desarrollo de lenguaje, coordinación motriz y conocimiento del cuerpo en preescolaresDocument6 pagesPlanificación de actividades para el desarrollo de lenguaje, coordinación motriz y conocimiento del cuerpo en preescolaresErika IñiguezNo ratings yet
- Manual Territory 2022 01Document172 pagesManual Territory 2022 01Carlos ChiesaNo ratings yet
- Autoevaluación N°1 - SIM 2 2Document4 pagesAutoevaluación N°1 - SIM 2 2MARION SHIRLEY FERNANDEZ CALDERONNo ratings yet
- Formulario de Áreas y Volúmenes. MatemáticasDocument3 pagesFormulario de Áreas y Volúmenes. MatemáticasdubanNo ratings yet
- Actividad 4Document8 pagesActividad 4Luz Angela Henao NavarroNo ratings yet
- Todas Las Diapositivas de Control de ProcesosDocument154 pagesTodas Las Diapositivas de Control de ProcesosJohnny SalazarNo ratings yet
- Mascarillas Caseras para El CabelloDocument9 pagesMascarillas Caseras para El CabelloGiselaSauceNo ratings yet
- FMS Practica N1Document50 pagesFMS Practica N1Cristian AndrangoNo ratings yet
- Funcionamiento Motor AsincronoDocument3 pagesFuncionamiento Motor AsincronoRoss OlejuaNo ratings yet
- Amor infinitoDocument18 pagesAmor infinitoCarlosSánchezAlayodeEnriquezNo ratings yet
- Ángulo de torsión y esfuerzos cortantes en un árbol de transmisión compuestoDocument21 pagesÁngulo de torsión y esfuerzos cortantes en un árbol de transmisión compuestoOlvera Lona JesúsNo ratings yet
- Capacitores Fijos y VariablesDocument5 pagesCapacitores Fijos y VariablesAlberto Lencinas100% (2)
- Vino de Postre y Sus Posibles MaridajesDocument55 pagesVino de Postre y Sus Posibles MaridajesEmmanuel BonninNo ratings yet
- Practica Calificada n1 de Mecanicanica de Fluidos IIDocument7 pagesPractica Calificada n1 de Mecanicanica de Fluidos IImanuel ssjNo ratings yet