You might also like
- DMT Plants List NeuroSoupDocument13 pagesDMT Plants List NeuroSoupKyle Burris100% (1)
- DMT Extraction Overview - The Basic ProcessDocument1 pageDMT Extraction Overview - The Basic ProcessAxel Gómez-Ortigoza100% (1)
- Through the Gateway of the Heart, Second Edition: Accounts and Experiences with MDMA and other Empathogenic SubstancesFrom EverandThrough the Gateway of the Heart, Second Edition: Accounts and Experiences with MDMA and other Empathogenic SubstancesNo ratings yet
- DMT PaperDocument12 pagesDMT PaperDRCSBoivin100% (1)
- DMT How & Why To Get Off by Gracie & ZarkovDocument6 pagesDMT How & Why To Get Off by Gracie & Zarkovread2012No ratings yet
- Cutter Wheel - H1140Document4 pagesCutter Wheel - H1140Sebastián Fernando Canul Mendez100% (2)
- ABB 3HAC050988 AM Arc and Arc Sensor RW 6-En PDFDocument238 pagesABB 3HAC050988 AM Arc and Arc Sensor RW 6-En PDForefat1No ratings yet
- Max Ion: The cyb/CTM Maximum Yield' ATB Salt TekDocument7 pagesMax Ion: The cyb/CTM Maximum Yield' ATB Salt Tekzack100% (1)
- DMTDocument7 pagesDMTcplowman230% (2)
- ACACIA SIMPLEX: A GUIDE TO EXTRACTING DMTDocument10 pagesACACIA SIMPLEX: A GUIDE TO EXTRACTING DMTrockers91No ratings yet
- Estrazione Di DMT Da Materiale VegetaleDocument30 pagesEstrazione Di DMT Da Materiale VegetaleMossad NewsNo ratings yet
- Q21Q21 Vinegar LimeDocument13 pagesQ21Q21 Vinegar LimesoftmunchieNo ratings yet
- Abn ExtractionDocument2 pagesAbn Extractionuttamkumar_mondol_5No ratings yet
- CENTRAL ANALGETICS Analgesics Chemistry Pharmacology of Drugs Daniel Lednicer 1982Document236 pagesCENTRAL ANALGETICS Analgesics Chemistry Pharmacology of Drugs Daniel Lednicer 1982BASKAR.LNo ratings yet
- How To ChangaDocument7 pagesHow To ChangaFlorianLiest100% (1)
- Novel Psychoactive Substances: Classification, Pharmacology and ToxicologyFrom EverandNovel Psychoactive Substances: Classification, Pharmacology and ToxicologyPaul I. DarganRating: 5 out of 5 stars5/5 (2)
- r32 - Rb26dett Ecu PinoutDocument1 pager32 - Rb26dett Ecu PinouttospotNo ratings yet
- McKerrall HallucinogensDocument13 pagesMcKerrall HallucinogensBrooke Rodriguez100% (1)
- The Psychopharmacology of HallucinogensFrom EverandThe Psychopharmacology of HallucinogensRichard C. StillmanNo ratings yet
- MAKING DMT GUIDE: The essential guide to master the art of making DMT the spirit molecule at home in a simplified wayFrom EverandMAKING DMT GUIDE: The essential guide to master the art of making DMT the spirit molecule at home in a simplified wayNo ratings yet
- MDMA (Ecstasy) - Final PresentationDocument12 pagesMDMA (Ecstasy) - Final PresentationNat Focha100% (1)
- MDMA Research PaperDocument7 pagesMDMA Research PaperKyle MooreNo ratings yet
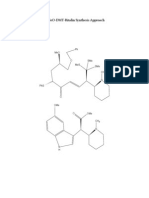
- 5 MeO DMT Ritalin Synthesis ApproachDocument5 pages5 MeO DMT Ritalin Synthesis ApproachCarlosHarmesNo ratings yet
- Erowid Newsletter17 PDFDocument28 pagesErowid Newsletter17 PDFRichard Portilloo100% (1)
- Psychedelic Book SuggestionsDocument13 pagesPsychedelic Book SuggestionsTomáš Kovačík50% (2)
- Extracción-DMT From Phragmites Australis - The Common ReedDocument9 pagesExtracción-DMT From Phragmites Australis - The Common Reedjona_19_02No ratings yet
- Shulgin Labbook3 SearchableDocument121 pagesShulgin Labbook3 Searchableoidane100% (3)
- Sasha Shulgin's 81st Birthday LimericksDocument62 pagesSasha Shulgin's 81st Birthday Limericksbigblue99100% (1)
- DMT (Dimethyltryptamine) : How To Use DMTDocument8 pagesDMT (Dimethyltryptamine) : How To Use DMTGeorge Valles100% (1)
- The Shulgin Pharmacology Labbooks: Book 1Document223 pagesThe Shulgin Pharmacology Labbooks: Book 1HoorayFrisbeeHead75% (4)
- The History of EcstasyDocument24 pagesThe History of Ecstasyshakahs204386% (7)
- MDMA ChemistryDocument15 pagesMDMA ChemistryJared FeldmanNo ratings yet
- Pharmacology Notes 01 Alexander ShulginDocument238 pagesPharmacology Notes 01 Alexander ShulginPedro Mendonca100% (3)
- 5meo SynthesisDocument2 pages5meo SynthesisLuis López PiñeresNo ratings yet
- Everything You Want To Know About RecrystallizationDocument3 pagesEverything You Want To Know About Recrystallizationrana_mohamed558100% (2)
- Erowid Newsletter12 PDFDocument28 pagesErowid Newsletter12 PDFRichard PortillooNo ratings yet
- SubstanceDocument11 pagesSubstancejrewingheroNo ratings yet
- MDMA For Anxiety DisordersDocument3 pagesMDMA For Anxiety DisordersSam JyotishiNo ratings yet
- Personality changes linked to psychedelicsDocument50 pagesPersonality changes linked to psychedelicslgessaNo ratings yet
- Rowid Xtracts: "In History There Are No Real Beginnings."Document28 pagesRowid Xtracts: "In History There Are No Real Beginnings."Richard Portilloo100% (1)
- Alkaloids Reported As (MG) Per (100g) Raw Dried Plant, andDocument1 pageAlkaloids Reported As (MG) Per (100g) Raw Dried Plant, andDBos1100% (2)
- Liquid-Liquid Extraction Experiment (LLEDocument5 pagesLiquid-Liquid Extraction Experiment (LLEMuhamad Baihakhi Shamsudin100% (1)
- The Entheogen Review׃ Vol. 16, No. 2 (2008)Document84 pagesThe Entheogen Review׃ Vol. 16, No. 2 (2008)HoorayFrisbeeHeadNo ratings yet
- Lyttle 1988Document14 pagesLyttle 1988Hamilton MorrisNo ratings yet
- An MDMA Neurochemistry SlideshowDocument18 pagesAn MDMA Neurochemistry SlideshowCraig Ohlsen100% (1)
- Ketamine - New TricksDocument9 pagesKetamine - New TricksIftach HuebnerNo ratings yet
- Ayahuasca - Entheogenic Education Public PolicyDocument348 pagesAyahuasca - Entheogenic Education Public Policysi1mas1es1No ratings yet
- Psychotria Viridis A Botanical Source of DimethyltryptamineDocument4 pagesPsychotria Viridis A Botanical Source of DimethyltryptaminetysonNo ratings yet
- Psychedelic Review, Vol. I, No. 4, 1964Document65 pagesPsychedelic Review, Vol. I, No. 4, 1964channel viewNo ratings yet
- Ben Sessa From Sacred Plants To Psychotherapy PDFDocument12 pagesBen Sessa From Sacred Plants To Psychotherapy PDFJim BennyNo ratings yet
- A New Species of Blue PoppyDocument6 pagesA New Species of Blue PoppyBhutan CanadaNo ratings yet
- Dimethyltryptamine (DMT)Document6 pagesDimethyltryptamine (DMT)Anonymous 6SykLi9No ratings yet
- DMT - Erowid Exp - 'Laboratory Synthesis'Document3 pagesDMT - Erowid Exp - 'Laboratory Synthesis'johannes karcher100% (1)
- Importance of Psychedelics by Terence McKennaDocument7 pagesImportance of Psychedelics by Terence McKennaread2012100% (1)
- The Untoward Effects of Drugs - A Pharmacological and Clinical ManualFrom EverandThe Untoward Effects of Drugs - A Pharmacological and Clinical ManualNo ratings yet
- Advances in Endogenous and Exogenous Opioids: Proceedings of the International Narcotic Research Conference (Satellite Symposium of the 8th International Congress of Pharmacology) Held in Kyoto, Japan on July 26–30, 1981From EverandAdvances in Endogenous and Exogenous Opioids: Proceedings of the International Narcotic Research Conference (Satellite Symposium of the 8th International Congress of Pharmacology) Held in Kyoto, Japan on July 26–30, 1981Hiroshi TakagiNo ratings yet
- Opioid Peptides: Biology, Chemistry, and Genetics: The Peptides: Analysis, Synthesis, Biology, Vol. 6From EverandOpioid Peptides: Biology, Chemistry, and Genetics: The Peptides: Analysis, Synthesis, Biology, Vol. 6No ratings yet
- Dopaminergic Ergot Derivatives and Motor Function: Proceedings of an International Symposium Held in the Wenner-Gren Center, Stockholm, July 24-25, 1978From EverandDopaminergic Ergot Derivatives and Motor Function: Proceedings of an International Symposium Held in the Wenner-Gren Center, Stockholm, July 24-25, 1978Kjell FuxeNo ratings yet
- 2005 DMT Jacob PrestiDocument8 pages2005 DMT Jacob PrestiМакс ПейнNo ratings yet
- BS en 12201 5 2011Document20 pagesBS en 12201 5 2011fatjon31100% (4)
- Philip Rance EAH Philo of ByzantiumDocument3 pagesPhilip Rance EAH Philo of ByzantiumstoliNo ratings yet
- Arta Fridei Kahlo A Fost Intotdeauna o ReactieDocument13 pagesArta Fridei Kahlo A Fost Intotdeauna o ReactieAlta DaianNo ratings yet
- Ebrosur Silk Town PDFDocument28 pagesEbrosur Silk Town PDFDausNo ratings yet
- Screenshot 2023-01-03 at 9.25.34 AM PDFDocument109 pagesScreenshot 2023-01-03 at 9.25.34 AM PDFAzri ZakwanNo ratings yet
- Products and Services For Consumers: True / False QuestionsDocument65 pagesProducts and Services For Consumers: True / False QuestionsThúy HiềnNo ratings yet
- Tipologie Break in OutDocument17 pagesTipologie Break in OutGiorgio FantauzziNo ratings yet
- Rincon Dueling RigbyDocument5 pagesRincon Dueling Rigbytootalldean100% (1)
- Orientation Report PDFDocument13 pagesOrientation Report PDFRiaz RasoolNo ratings yet
- Unit 4.2Document45 pagesUnit 4.2Gundrathi Narendra GoudNo ratings yet
- H. Bateman, A. Erdélyi Et Al. - Higher Transcendental Functions 3 (1955, McGraw-Hill)Document310 pagesH. Bateman, A. Erdélyi Et Al. - Higher Transcendental Functions 3 (1955, McGraw-Hill)ITALO HERRERA MOYANo ratings yet
- CV1000 CV2000 CV3000 Ice Cube Machines SpecsDocument12 pagesCV1000 CV2000 CV3000 Ice Cube Machines SpecsAriel Godoy100% (1)
- BMW Mini COoper Installation InstructionsDocument1 pageBMW Mini COoper Installation InstructionsEdiJonNo ratings yet
- Weber Grills - FinalDocument12 pagesWeber Grills - FinalDIVYANSHU SHEKHARNo ratings yet
- Tap Yourself FreeDocument134 pagesTap Yourself Freenguyenhavn100% (2)
- Medium Strength High Conductivity MaterialsDocument37 pagesMedium Strength High Conductivity MaterialsNut AssanaiNo ratings yet
- QUESTION A2 and A3 Examples PDFDocument24 pagesQUESTION A2 and A3 Examples PDFDemian VeNo ratings yet
- Measuring and calculating dimensions for pipes, plates, cylinders and moreDocument100 pagesMeasuring and calculating dimensions for pipes, plates, cylinders and moreGarcia MaybelleNo ratings yet
- 3 Variable Cramers Rule PDFDocument4 pages3 Variable Cramers Rule PDFElton DiocsonNo ratings yet
- Progressing Cavity Pump Overhaul GuideDocument5 pagesProgressing Cavity Pump Overhaul Guidesdsds-54No ratings yet
- Poultry DiseasesDocument5 pagesPoultry DiseasesAnjum IslamNo ratings yet
- 2021 - Tet Purchase Behavior Report - INFOCUSDocument15 pages2021 - Tet Purchase Behavior Report - INFOCUSGame AccountNo ratings yet
- GMsetDocument8 pagesGMsetdilo001No ratings yet
- Ch3 XII SolutionsDocument12 pagesCh3 XII SolutionsSaish NaikNo ratings yet
- Solid Waste ManagementDocument26 pagesSolid Waste ManagementPamela MendozaNo ratings yet
- SDE1 V1 G2 H18 L P2 M8 - SpecificationsDocument1 pageSDE1 V1 G2 H18 L P2 M8 - SpecificationsCleverson SoaresNo ratings yet
- Belden CatalogDocument24 pagesBelden CatalogMani MaranNo ratings yet