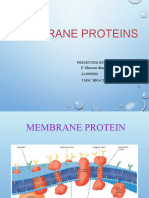
You might also like
- C9e Answers Active Reading 07Document7 pagesC9e Answers Active Reading 07Daniel DHizzy Hizgilov83% (18)
- The Cellular Level of OrganizationDocument7 pagesThe Cellular Level of OrganizationPearl MirandaNo ratings yet
- Biology - Study and Revision Guide - Andrew Davis and C. J. Clegg - Second Edition - Hodder 2017Document537 pagesBiology - Study and Revision Guide - Andrew Davis and C. J. Clegg - Second Edition - Hodder 2017ariel camposNo ratings yet
- Nelson Biology Answers 1Document2 pagesNelson Biology Answers 1ritaNo ratings yet
- Biological Cell MembranesDocument44 pagesBiological Cell MembranesMohammed ShaikNo ratings yet
- Membrane Proteins2Document29 pagesMembrane Proteins2spdharanimaran001No ratings yet
- Kuliah MembranDocument61 pagesKuliah Membrannikmah nuur rochmahNo ratings yet
- BIO 315. NotesDocument42 pagesBIO 315. NotesKevin KipropNo ratings yet
- Summary Essential Cell Biology Chapters 4678121516 1718 and 20Document36 pagesSummary Essential Cell Biology Chapters 4678121516 1718 and 20mohamed arbi taifNo ratings yet
- Study Exercise Biology 10-12 97.03Document7 pagesStudy Exercise Biology 10-12 97.03Fabiola Vania FeliciaNo ratings yet
- N-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andDocument9 pagesN-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andHeisenbergNo ratings yet
- The Cell MembraneDocument36 pagesThe Cell Membraneanulorance98No ratings yet
- Protein TransportDocument13 pagesProtein TransportvmshanesNo ratings yet
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- If You Understand How Proteins WorkDocument4 pagesIf You Understand How Proteins WorkMah Song WeiNo ratings yet
- LO Week 7Document22 pagesLO Week 7Matthew ChristopherNo ratings yet
- Membrane Proteins: Functions & Types in 40 CharactersDocument7 pagesMembrane Proteins: Functions & Types in 40 CharactersUsman HashmiNo ratings yet
- Fluid Mosaic Paper PDFDocument0 pagesFluid Mosaic Paper PDFDina Kharida100% (1)
- Concise Histology Gartner, Leslie PDFDocument478 pagesConcise Histology Gartner, Leslie PDFLexsshNo ratings yet
- Cell Biology Lec 5-2017Document41 pagesCell Biology Lec 5-2017Nitesh SardarNo ratings yet
- Assignment For Membrane StructureDocument3 pagesAssignment For Membrane Structurejane austin lynn rebancosNo ratings yet
- Cell Membrane Transport ActivityDocument7 pagesCell Membrane Transport ActivityoctavianojanineNo ratings yet
- Cell Membranes and SignalingDocument7 pagesCell Membranes and Signalingbasmala6545No ratings yet
- ANSWEREEDocument3 pagesANSWEREERaxilla MaxillaNo ratings yet
- Research Paper On Plasma MembraneDocument4 pagesResearch Paper On Plasma Membranevvgnzdbkf100% (1)
- Cell Membrane SummaryDocument1 pageCell Membrane SummaryzafNo ratings yet
- Zoology Lecture 4Document18 pagesZoology Lecture 4Stephen OcheaNo ratings yet
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- Chapter 7Document4 pagesChapter 7Astrii LyNo ratings yet
- Chapter 4-Jnu Biomembranes-1 - 複本Document48 pagesChapter 4-Jnu Biomembranes-1 - 複本Wai Kwong ChiuNo ratings yet
- Components of Plasma Membranes: Learning ObjectivesDocument14 pagesComponents of Plasma Membranes: Learning ObjectivesAlthea Junsan BernabeNo ratings yet
- Cell Organelles and Their FunctionsDocument8 pagesCell Organelles and Their FunctionsPrincess Angie GonzalesNo ratings yet
- 16.plasma membr-WPS OfficeDocument4 pages16.plasma membr-WPS OfficeMohamed RizwanNo ratings yet
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowNo ratings yet
- SF - MD.0145.T.22 Ally Omary MwambelaDocument10 pagesSF - MD.0145.T.22 Ally Omary MwambelaAlly OmaryNo ratings yet
- Functional and Structural Properties of Natural BiomembranesDocument7 pagesFunctional and Structural Properties of Natural BiomembranesAhsan RazaNo ratings yet
- Proteins: Structure & FunctionsDocument15 pagesProteins: Structure & FunctionsAbhinav KumarNo ratings yet
- Module 10 ACTDocument5 pagesModule 10 ACTLeighNo ratings yet
- Transport of Substances Through Cell MembranesDocument34 pagesTransport of Substances Through Cell MembranesSanchezNo ratings yet
- CSIR Life Science Sample Cell BiologyDocument17 pagesCSIR Life Science Sample Cell Biologygamer zoneNo ratings yet
- Plasma Membrane in DetailsDocument24 pagesPlasma Membrane in DetailsSyeda Kaneez Mohammadi MohammadiNo ratings yet
- Unit 2 ReviewerDocument4 pagesUnit 2 Revieweraaleah moscaNo ratings yet
- Chapter 1doc 3Document4 pagesChapter 1doc 3Enju ChipepeNo ratings yet
- CELL DIFFERENTIATION AND ORGANELLESDocument13 pagesCELL DIFFERENTIATION AND ORGANELLESKyleBernalÜNo ratings yet
- 2structure and Functions of Cell MembraneDocument19 pages2structure and Functions of Cell MembraneedriansamaNo ratings yet
- Plasma Membrane Research PapersDocument6 pagesPlasma Membrane Research Papersaflbasnka100% (1)
- Mammalian Histology AssignmentDocument9 pagesMammalian Histology AssignmentSana Sultana100% (1)
- Semipermeable MembraneDocument1 pageSemipermeable MembranedeniseeeeeeNo ratings yet
- Struktur Dan Fungsi Protein: Joko MarwotoDocument66 pagesStruktur Dan Fungsi Protein: Joko MarwotoBilly PeterNo ratings yet
- Histo Chapter 2Document2 pagesHisto Chapter 2AnnNo ratings yet
- IB HL Biology Notes Cell MembranesDocument12 pagesIB HL Biology Notes Cell MembranesayushfmNo ratings yet
- Chapter7 - Membrane Structure and FunctionDocument13 pagesChapter7 - Membrane Structure and FunctionKrizziane Ivy CuajotorNo ratings yet
- Assignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3ADocument9 pagesAssignment For Membrane Structure: Ane Austin Lynn N. Rebancos Bsed-Science 3Ajane austin lynn rebancosNo ratings yet
- PHYSIOLOGYDocument16 pagesPHYSIOLOGYIqtader hoque khan AkibNo ratings yet
- Fluid Mosaic ModelDocument3 pagesFluid Mosaic Modelmariam manzoorNo ratings yet
- 9.1C: Types of Receptors: Learning ObjectivesDocument3 pages9.1C: Types of Receptors: Learning Objectivesstalker akoNo ratings yet
- Cell MembraneDocument15 pagesCell MembraneAyesha SaleemNo ratings yet
- Lipid Traffic Floppy Drives and A SuperhighwayDocument12 pagesLipid Traffic Floppy Drives and A SuperhighwayJohan AlarcónNo ratings yet
- NRS1102: Anatomy and Physiology (Lecture) 1 Semester 2020-2021 Ejfmania DMD, RN, MANDocument14 pagesNRS1102: Anatomy and Physiology (Lecture) 1 Semester 2020-2021 Ejfmania DMD, RN, MANEllen Mynelle MabulacNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture Outlinehaha_le12No ratings yet
- Mechanisms and Regulation of Carbohydrate Transport in BacteriaFrom EverandMechanisms and Regulation of Carbohydrate Transport in BacteriaNo ratings yet
- Chord Diagram With Time SliderDocument4 pagesChord Diagram With Time SliderbahlasaNo ratings yet
- Diff GeoDocument252 pagesDiff GeoKirti Deo Mishra100% (1)
- B&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. CDocument4 pagesB&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. CbahlasaNo ratings yet
- B&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. ADocument5 pagesB&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. AbahlasaNo ratings yet
- B&B's Medical Physiology. 2. Functional Organization of The Cell - IntroDocument1 pageB&B's Medical Physiology. 2. Functional Organization of The Cell - IntrobahlasaNo ratings yet
- B&B's Medical Physiology. 2. Functional Organization of The Cell - IntroDocument1 pageB&B's Medical Physiology. 2. Functional Organization of The Cell - IntrobahlasaNo ratings yet
- B&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. (Intro)Document1 pageB&B Medical Physiology - II. Physiology of Cells and Molecules. 3. Physiology of Membranes. (Intro)bahlasaNo ratings yet
- B&B Medical Physiology - II. Physiology of Cells and Molecules. 2. Functional Organization of The Cell. ADocument9 pagesB&B Medical Physiology - II. Physiology of Cells and Molecules. 2. Functional Organization of The Cell. AbahlasaNo ratings yet
- Mechanistic Insights of Formulation Approaches For The Treatment of Nail Infection - Conventional and Novel Drug Delivery ApproachesDocument12 pagesMechanistic Insights of Formulation Approaches For The Treatment of Nail Infection - Conventional and Novel Drug Delivery ApproachessovalaxNo ratings yet
- Cell Membrane Transport (Compatibility Mode)Document12 pagesCell Membrane Transport (Compatibility Mode)Kishore Singh BasnetNo ratings yet
- Lec 8Document34 pagesLec 8Shraddha JaiswalNo ratings yet
- Semester Exam Study Guide KEYDocument17 pagesSemester Exam Study Guide KEYOmar CamposNo ratings yet
- RANKUP NEET (2019) DAY -1 (DPP) Geometrical Optics and Stoichiometry QuestionsDocument32 pagesRANKUP NEET (2019) DAY -1 (DPP) Geometrical Optics and Stoichiometry QuestionsYash bhartiNo ratings yet
- Terahertz Radiation Effects and Biological ApplicationsDocument11 pagesTerahertz Radiation Effects and Biological Applicationsjiwei tianNo ratings yet
- Plasma MembraneDocument40 pagesPlasma Membraneshaffan ula prasetyoNo ratings yet
- University of Gondar College of Medicine and Health Sciences Department of Medical PhysiologyDocument202 pagesUniversity of Gondar College of Medicine and Health Sciences Department of Medical PhysiologyGifti DemisseNo ratings yet
- Original Pdfbiology Exploring The Diversity of Life 4th 1 3 by Peter Russell PDFDocument41 pagesOriginal Pdfbiology Exploring The Diversity of Life 4th 1 3 by Peter Russell PDFbonnie.eaton513100% (34)
- (Carbon Nanostructures) Gil Gonçalves, Paula Marques, Mercedes Vila (Eds.) - Graphene-Based Materials in Health and Environment - New Paradigms-Springer International Publishing (2016)Document358 pages(Carbon Nanostructures) Gil Gonçalves, Paula Marques, Mercedes Vila (Eds.) - Graphene-Based Materials in Health and Environment - New Paradigms-Springer International Publishing (2016)lonelystar19No ratings yet
- BIO Ch03 MC eDocument35 pagesBIO Ch03 MC ekenchan0810.kcNo ratings yet
- Detergent Handbook PDFDocument16 pagesDetergent Handbook PDFfahmyaziz88% (16)
- Code RCH 2003Document23 pagesCode RCH 2003Rhenso Victor Albites CondoriNo ratings yet
- Encapsulation PPT AkashDocument18 pagesEncapsulation PPT AkashAkash PandayNo ratings yet
- Vesicular Drug Delivery System: A Novel Approach: September 2014Document15 pagesVesicular Drug Delivery System: A Novel Approach: September 2014Hely PatelNo ratings yet
- Mitochondria and ChloroplastDocument71 pagesMitochondria and ChloroplastRifai Alfarabi100% (1)
- Trends in Drug ResearchDocument186 pagesTrends in Drug ResearchEvsevios HadjicostasNo ratings yet
- Cell Membrane Structure and Function ExplainedDocument10 pagesCell Membrane Structure and Function ExplainedkyleeebooggNo ratings yet
- Lecture 2 Membrane Transport & Cell Signaling - QuestionsDocument6 pagesLecture 2 Membrane Transport & Cell Signaling - Questions中华雅思王No ratings yet
- Cell Theory and Biochemical Aspects of Cell MembraneDocument39 pagesCell Theory and Biochemical Aspects of Cell MembraneHaroon BadarNo ratings yet
- Biology PAKMCQsDocument290 pagesBiology PAKMCQsMashooque AliNo ratings yet
- Terconazole Proniosomal Gels Effect of Different Formulation Factors Physicochemical and Microbiological Evaluation WJAZDocument6 pagesTerconazole Proniosomal Gels Effect of Different Formulation Factors Physicochemical and Microbiological Evaluation WJAZWiri Resky AmaliaNo ratings yet
- A Novel Approach For Iron DeficiencyDocument15 pagesA Novel Approach For Iron DeficiencyMahyar MehrNo ratings yet
- Chapter - 08 - Lipids Membranes-TestDocument10 pagesChapter - 08 - Lipids Membranes-Testendang dian lestariNo ratings yet
- Cell Membrane Structure and Function: The Fluid Mosaic ModelDocument75 pagesCell Membrane Structure and Function: The Fluid Mosaic Modelvinay chaudhary100% (1)
- MCQ Cell Structure and FunctionDocument14 pagesMCQ Cell Structure and FunctionVirendra Joshi80% (5)
- Lipids Exam Study GuideDocument17 pagesLipids Exam Study GuideGreg ZhangNo ratings yet
- Perspectives: Coexistence of Passive and Carrier-Mediated Processes in Drug TransportDocument18 pagesPerspectives: Coexistence of Passive and Carrier-Mediated Processes in Drug TransportzaynmalikNo ratings yet