You might also like
- Thesis About YemenDocument488 pagesThesis About Yemenmarwan98No ratings yet
- A Protocol For Extraction and Purification of High-Quality and Quantity Bacterial DNA Applicable For Genome Sequencing: A Modified Version of The Marmur Procedure.Document6 pagesA Protocol For Extraction and Purification of High-Quality and Quantity Bacterial DNA Applicable For Genome Sequencing: A Modified Version of The Marmur Procedure.marwan98No ratings yet
- pUC 19 Ligation PDFDocument6 pagespUC 19 Ligation PDFmarwan98No ratings yet
- Miswak As An Alternative To The Modern Toothbrush in Preventing Oral DiseasesDocument33 pagesMiswak As An Alternative To The Modern Toothbrush in Preventing Oral Diseasesmarwan98No ratings yet
- 09 UV Absorption PDFDocument4 pages09 UV Absorption PDFyongqeeNo ratings yet
- Chemical and biologically active components of the toothbrush tree (Salvadora persicaDocument6 pagesChemical and biologically active components of the toothbrush tree (Salvadora persicamarwan98No ratings yet
- Oral Squamous Cell Carcinoma OverviewDocument8 pagesOral Squamous Cell Carcinoma Overviewmarwan98No ratings yet
- The Vegetation of SocotraDocument21 pagesThe Vegetation of Socotramarwan98No ratings yet
- 1 PBDocument9 pages1 PBmarwan98No ratings yet
- Gades Poster November 2009 - MarwanDocument1 pageGades Poster November 2009 - Marwanmarwan98No ratings yet
- ﻣﺸﺮﻭﻉ ﺍﻟﺪﺳﺘﻮﺭ ﺍﳉﺪﻳﺪ ﻟﻠﺪﻭﻟﺔ ﺍﳌﺼﺮﻳﺔ ﺍﳊﺪﻳﺜﺔDocument49 pagesﻣﺸﺮﻭﻉ ﺍﻟﺪﺳﺘﻮﺭ ﺍﳉﺪﻳﺪ ﻟﻠﺪﻭﻟﺔ ﺍﳌﺼﺮﻳﺔ ﺍﳊﺪﻳﺜﺔgaa5No ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Isolation and Identification Ort From ChickenDocument14 pagesIsolation and Identification Ort From Chickenthanh ba matNo ratings yet
- 05 Introduction To Plant PathologyDocument20 pages05 Introduction To Plant PathologyirwanpaluNo ratings yet
- Microbiology 205 Written Exam 1Document2 pagesMicrobiology 205 Written Exam 1marieeepNo ratings yet
- Hepatitis G Virus (HGV) at GB Virus C (GBV-C)Document18 pagesHepatitis G Virus (HGV) at GB Virus C (GBV-C)denimm1991No ratings yet
- Medical Micro MCQsDocument26 pagesMedical Micro MCQsAhmad RajoubNo ratings yet
- Historical OVERVIEW On Blood BankingDocument4 pagesHistorical OVERVIEW On Blood BankingSogan, MaureenNo ratings yet
- Isolation and Identification of Avibacterium Paragallinarum From Layer Chickens in Gazipur, BangladeshDocument3 pagesIsolation and Identification of Avibacterium Paragallinarum From Layer Chickens in Gazipur, BangladeshMd. Ehsanul Haque100% (1)
- Blood ComponentsDocument44 pagesBlood ComponentsKrisha VittoNo ratings yet
- Bacterial and viral intestinal infectionsDocument1,786 pagesBacterial and viral intestinal infectionsalvin bramantyoNo ratings yet
- Blood Grouping ReagentsDocument7 pagesBlood Grouping ReagentsDominic EmerencianaNo ratings yet
- Sodium, Blood ,: Test Name Result Biological Ref. Interval MethodDocument3 pagesSodium, Blood ,: Test Name Result Biological Ref. Interval MethodPantha BiswasNo ratings yet
- Bacteria vs Virus: Key Differences and How Pathogenic Bacteria Harm the Human BodyDocument11 pagesBacteria vs Virus: Key Differences and How Pathogenic Bacteria Harm the Human BodySakib Rahman DiptoNo ratings yet
- Lab ReportDocument10 pagesLab ReportJeevikaGoyal100% (1)
- Blood Type Genetics 40Document3 pagesBlood Type Genetics 40Gary Garlan100% (1)
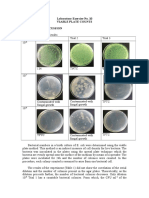
- Laboratory Exercise No. 10 Viable Plate Counts Results and DiscussionDocument3 pagesLaboratory Exercise No. 10 Viable Plate Counts Results and Discussionvanessa olga100% (1)
- Compatibility Testing For Blood TransfusionDocument40 pagesCompatibility Testing For Blood TransfusionMunish Dogra100% (4)
- Plateletcrit As A Screening Tool For Detection of Platelet Quantitative DisordersDocument5 pagesPlateletcrit As A Screening Tool For Detection of Platelet Quantitative DisordersStephanie PattonNo ratings yet
- Ra 7719 Blood Bank MT-B & C 8Document7 pagesRa 7719 Blood Bank MT-B & C 8Fait Hee100% (1)
- 8.29.06 Staphlococcal BacteremiaDocument41 pages8.29.06 Staphlococcal BacteremiaVaibhav KaroliyaNo ratings yet
- Laboratorio Clinico Parque Escorial: Patient Number Birthdate SexDocument1 pageLaboratorio Clinico Parque Escorial: Patient Number Birthdate SexCarol Melissa AquinoNo ratings yet
- Transfusion MedicineDocument4 pagesTransfusion MedicineMaikka IlaganNo ratings yet
- 2 Raos ISBT Immunohematology WP Case Study 1 FINAL PDFDocument31 pages2 Raos ISBT Immunohematology WP Case Study 1 FINAL PDFdrtsotlhe mogolegwaNo ratings yet
- Bacteria: Small Living Things That Can Be Found Almost EverywhereDocument4 pagesBacteria: Small Living Things That Can Be Found Almost EverywhereIsmilNo ratings yet
- ICSE Biology Economic Significance of MicroorganismsDocument42 pagesICSE Biology Economic Significance of MicroorganismsDebasish KunduNo ratings yet
- US Army Medical Course MD0856-200 - BacteriologyDocument267 pagesUS Army Medical Course MD0856-200 - BacteriologyGeorges100% (2)
- Medical Mnemonics For MicroorgDocument4 pagesMedical Mnemonics For Microorgkenz18096No ratings yet
- Blood Grouping Brochure 2013 - Priced PDFDocument8 pagesBlood Grouping Brochure 2013 - Priced PDFmwenyeweNo ratings yet
- Microorganisms: Friend and Foe Class 8 Extra Questions Science ChapterDocument10 pagesMicroorganisms: Friend and Foe Class 8 Extra Questions Science ChapterManjula ManjulaNo ratings yet
- Lab Blood TypesDocument3 pagesLab Blood TypesTrizia M MendezNo ratings yet
- CHC Jawa Hub: Jawa, Rewa, Madhya Pradesh Rewa Madhya Pradesh - 486223 Phone No.Document2 pagesCHC Jawa Hub: Jawa, Rewa, Madhya Pradesh Rewa Madhya Pradesh - 486223 Phone No.MAHESH GAUTAMNo ratings yet