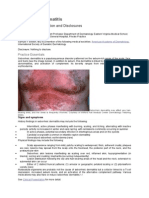
You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- How Vaccines Can Damage Your Brain, Russell BlaylockDocument9 pagesHow Vaccines Can Damage Your Brain, Russell BlaylockCherie RaymondNo ratings yet
- Overdosed Babies:: Are Multiple Vaccines Safe?Document0 pagesOverdosed Babies:: Are Multiple Vaccines Safe?AprianaRohman100% (1)
- Verbal Probability Expressions in National Intelligence EstimatesDocument124 pagesVerbal Probability Expressions in National Intelligence EstimatesKristan J. Wheaton100% (4)
- EczemaDocument23 pagesEczemaPn Ziza AzwaNo ratings yet
- Health 8 Q3 Week 7-8 Module 1-6Document22 pagesHealth 8 Q3 Week 7-8 Module 1-6Richard CoderiasNo ratings yet
- Seborrheic Dermatitis EmedicineDocument10 pagesSeborrheic Dermatitis EmedicineIkram IkramNo ratings yet
- 06 Cross ImpactDocument19 pages06 Cross ImpactjaherrerarNo ratings yet
- Bronchial AsthmaDocument29 pagesBronchial AsthmaDrShaheen100% (5)
- Dermatologic Immunity PDFDocument391 pagesDermatologic Immunity PDFChino UnjbgNo ratings yet
- General Dermatology MCQsDocument197 pagesGeneral Dermatology MCQsDr.Tawheed95% (19)
- Immunoglobulin A (Iga)Document1 pageImmunoglobulin A (Iga)Lione MilanNo ratings yet
- NCP PEDIA TonsillitisDocument2 pagesNCP PEDIA Tonsillitisdeliejoyce100% (1)
- Powerpivot Dax Functions LongDocument5 pagesPowerpivot Dax Functions LongjaherrerarNo ratings yet
- Foreknowledge Issue 5Document22 pagesForeknowledge Issue 5jaherrerarNo ratings yet
- DSAIDE Package Tutorial: Interactive Exploration of Infectious Disease ModelsDocument9 pagesDSAIDE Package Tutorial: Interactive Exploration of Infectious Disease ModelsjaherrerarNo ratings yet
- Understanding LampDocument13 pagesUnderstanding Lampjaherrerar100% (1)
- Math Puts Ractopamine Economics in PerspectiveDocument3 pagesMath Puts Ractopamine Economics in PerspectivejaherrerarNo ratings yet
- Machine Milking of Ostfriesian and Lacaune Dairy Sheep: Udder Anatomy, Milk Ejection and Milking CharacteristicsDocument10 pagesMachine Milking of Ostfriesian and Lacaune Dairy Sheep: Udder Anatomy, Milk Ejection and Milking CharacteristicsjaherrerarNo ratings yet
- Vet - Res. 0928-4249 2008 39 2 ART0003Document14 pagesVet - Res. 0928-4249 2008 39 2 ART0003jaherrerarNo ratings yet
- 96Document169 pages96jaherrerarNo ratings yet
- Tables enDocument12 pagesTables enjaherrerarNo ratings yet
- Pork Sector in UruguayDocument13 pagesPork Sector in UruguayjaherrerarNo ratings yet
- Comparacion IndustriasDocument6 pagesComparacion IndustriasjaherrerarNo ratings yet
- Updating of The Correlation Between LpELISA Titers and Protection From Virus Challenge For The Assessment of The Potency of Polyvalent Aphtovirus Vaccines in ArgentinaDocument10 pagesUpdating of The Correlation Between LpELISA Titers and Protection From Virus Challenge For The Assessment of The Potency of Polyvalent Aphtovirus Vaccines in ArgentinajaherrerarNo ratings yet
- Vaccination Against Foot and Mouth Disease Reduces Virus Transmission in Groups of CalvesDocument8 pagesVaccination Against Foot and Mouth Disease Reduces Virus Transmission in Groups of CalvesjaherrerarNo ratings yet
- Use of Continuous Results To Compare ELISAs For The Detection of Antibodies To Non-Structural Proteins of Foot-And-mouth Disease VirusDocument10 pagesUse of Continuous Results To Compare ELISAs For The Detection of Antibodies To Non-Structural Proteins of Foot-And-mouth Disease VirusjaherrerarNo ratings yet
- Viral Infections and Bovine Mastitis - A ReviewDocument19 pagesViral Infections and Bovine Mastitis - A ReviewjaherrerarNo ratings yet
- Mra 11Document52 pagesMra 11jaherrerarNo ratings yet
- The Role of Small Ruminants in The Epidemiology and Transmission of Foot-And-Mouth DiseaseDocument8 pagesThe Role of Small Ruminants in The Epidemiology and Transmission of Foot-And-Mouth DiseasejaherrerarNo ratings yet
- The Tale of A Modern Animal Plague - Tracing The Evolutionary History and Determining The Time-Scale For Foot and Mouth Disease VirusDocument7 pagesThe Tale of A Modern Animal Plague - Tracing The Evolutionary History and Determining The Time-Scale For Foot and Mouth Disease VirusjaherrerarNo ratings yet
- Thermostable Foot-And-mouth Disease Virus As A Vaccine Candidate For Endemic Countries - A PerspectiveDocument3 pagesThermostable Foot-And-mouth Disease Virus As A Vaccine Candidate For Endemic Countries - A PerspectivejaherrerarNo ratings yet
- Vital Elements For The Successful Control of Foot-And-mouth Disease by VaccinationDocument7 pagesVital Elements For The Successful Control of Foot-And-mouth Disease by VaccinationjaherrerarNo ratings yet
- Vaccination of Pigs Two Weeks Before Infection Significantly Reduces Transmission of Foot-And-mouth Disease VirusDocument7 pagesVaccination of Pigs Two Weeks Before Infection Significantly Reduces Transmission of Foot-And-mouth Disease VirusjaherrerarNo ratings yet
- The Suitability of The Emergency' Foot-And-Mouth Disease Antigens Held by The International Vaccine Bank Within A Global ContextDocument11 pagesThe Suitability of The Emergency' Foot-And-Mouth Disease Antigens Held by The International Vaccine Bank Within A Global ContextjaherrerarNo ratings yet
- The Role of Infectious Aerosols in Disease Transmission in PigsDocument18 pagesThe Role of Infectious Aerosols in Disease Transmission in PigsjaherrerarNo ratings yet
- The NSP Immune Response of Vaccinated Animals After In-Field Exposure To FMDVDocument8 pagesThe NSP Immune Response of Vaccinated Animals After In-Field Exposure To FMDVjaherrerarNo ratings yet
- The Region Between The Two Polyprotein Initiation Codons of Foot-And-mouth Disease Virus Is Critical For Virulence in CattleDocument8 pagesThe Region Between The Two Polyprotein Initiation Codons of Foot-And-mouth Disease Virus Is Critical For Virulence in CattlejaherrerarNo ratings yet
- Vaccination Markers - Designing Unique Antigens To Be Added To Vaccines To Differentiate Between Natural Infection and VaccinationDocument3 pagesVaccination Markers - Designing Unique Antigens To Be Added To Vaccines To Differentiate Between Natural Infection and VaccinationjaherrerarNo ratings yet
- Foot-And-mouth Disease Vaccine Potency Testing - Determination and Statistical Validation of A Model Using A Serological ApproachDocument9 pagesFoot-And-mouth Disease Vaccine Potency Testing - Determination and Statistical Validation of A Model Using A Serological ApproachjaherrerarNo ratings yet
- Vaccination Against Foot-And-mouth Disease Virus Confers Complete Clinical Protection in 7 Days and Partial Protection in 4 Days - Use in Emergency Outbreak ResponseDocument8 pagesVaccination Against Foot-And-mouth Disease Virus Confers Complete Clinical Protection in 7 Days and Partial Protection in 4 Days - Use in Emergency Outbreak ResponsejaherrerarNo ratings yet
- Causes and Types of Cheilitis Inflammation of the LipsDocument25 pagesCauses and Types of Cheilitis Inflammation of the LipsFaiza LailiyahNo ratings yet
- Puskesmas Wanggudu Raya: Dinas Kesehatan Kabupaten Konawe UtaraDocument6 pagesPuskesmas Wanggudu Raya: Dinas Kesehatan Kabupaten Konawe UtaraicaNo ratings yet
- Feline PanleukopeniaDocument4 pagesFeline PanleukopeniaDebora BiantiiNo ratings yet
- Prevent Dengue: Learn Aedes Mosquito FactsDocument31 pagesPrevent Dengue: Learn Aedes Mosquito FactsreemarifkhanNo ratings yet
- BitesDocument6 pagesBitesAbubakr Hassan AhmadNo ratings yet
- Blood Bank Visit ReportDocument13 pagesBlood Bank Visit ReportsysiddhiyadavNo ratings yet
- Viral Hepatitis in ChildrenDocument15 pagesViral Hepatitis in ChildrenDr AnilNo ratings yet
- Antiobiotics Use Them Wisely 800 Passage and QuestionsDocument5 pagesAntiobiotics Use Them Wisely 800 Passage and Questionsdinh lanNo ratings yet
- National Action Plan - Viral Hepatitis Control Programmelowress - Reference File PDFDocument52 pagesNational Action Plan - Viral Hepatitis Control Programmelowress - Reference File PDFFranklin ThomasNo ratings yet
- WBC Differential CountDocument57 pagesWBC Differential CountSamantha IsabelNo ratings yet
- Nigella SativaDocument9 pagesNigella SativaoshinNo ratings yet
- Bacteriology. Virusology. Parasitology. - Exam 2021 - 20221 99 1Document70 pagesBacteriology. Virusology. Parasitology. - Exam 2021 - 20221 99 1Aron SlavescuNo ratings yet
- Histology of Tonsil and SpleenDocument27 pagesHistology of Tonsil and SpleenhusshamNo ratings yet
- Bicol University College of Nursing Protozoans ComparisonDocument3 pagesBicol University College of Nursing Protozoans ComparisonAbegail ListancoNo ratings yet
- BACTE (Quizlet)Document4 pagesBACTE (Quizlet)Allyssa AniNo ratings yet
- Autoimmune destruction of pancreatic beta cellsDocument12 pagesAutoimmune destruction of pancreatic beta cellsBea EscobarNo ratings yet
- 23 Avian Leukosis Complex 1Document14 pages23 Avian Leukosis Complex 1Aqib HussainNo ratings yet
- Master Question List For Covid-19 (Caused by Sars-Cov-2) : Dhs Science and TechnologyDocument26 pagesMaster Question List For Covid-19 (Caused by Sars-Cov-2) : Dhs Science and Technologybizonel84No ratings yet
- Virus Chart For Med MicroDocument2 pagesVirus Chart For Med MicroAndie RaczNo ratings yet
- Antigen-Antibody ReactionsDocument16 pagesAntigen-Antibody ReactionsArshdeep SinghNo ratings yet