You might also like
- Fisiología y Fisiopatología de Los Músculos de La EstáticaDocument24 pagesFisiología y Fisiopatología de Los Músculos de La EstáticaCristina Ortiz GuillénNo ratings yet
- Tema 7. Mecánica Muscular.Document10 pagesTema 7. Mecánica Muscular.Juan Ignacio Muñio BerdejoNo ratings yet
- Tema III.1 FuerzaDocument40 pagesTema III.1 FuerzaPABLO ENRÍQUEZ ENRÍQUEZNo ratings yet
- Biomecánica músculo estructuraDocument20 pagesBiomecánica músculo estructuraHumphrey Cruzado Catalan SandovalNo ratings yet
- Articulo MúsculoDocument13 pagesArticulo MúsculoJuan Camilo MenesesNo ratings yet
- La Flexibilidad.Document23 pagesLa Flexibilidad.LorenzoNo ratings yet
- Observaciones Sobre Los Distintos Tipos de Células o Fibras Musculares A Nivel Metabólico, Molecular y Su Rendimiento en La Actividad FísicaDocument10 pagesObservaciones Sobre Los Distintos Tipos de Células o Fibras Musculares A Nivel Metabólico, Molecular y Su Rendimiento en La Actividad FísicaLudmila CaseresNo ratings yet
- M1L3Document21 pagesM1L3Paola ANo ratings yet
- Biomecanica de MusculoDocument7 pagesBiomecanica de MusculoAnonymous 3hx8Y7LNo ratings yet
- Sistema Muscular - Wikipedia, La Enciclopedia LibreDocument20 pagesSistema Muscular - Wikipedia, La Enciclopedia LibreSteven RiveraNo ratings yet
- GD1-Entrenamiento PersonalizadoDocument33 pagesGD1-Entrenamiento PersonalizadoCriszehro ReviewerNo ratings yet
- Capítulo 1 Fisiología de La Contracción Muscular - APADocument18 pagesCapítulo 1 Fisiología de La Contracción Muscular - APAManuel Alejandro Osorio ArzapaloNo ratings yet
- ResumenDocument13 pagesResumenximena alvarezNo ratings yet
- Contraccion Muscular Final-2Document13 pagesContraccion Muscular Final-2Xochilth MairenaNo ratings yet
- Tema III.2 FlexibilidadDocument9 pagesTema III.2 FlexibilidadPABLO ENRÍQUEZ ENRÍQUEZNo ratings yet
- Sistema Muscular de Los Seres HumanosDocument3 pagesSistema Muscular de Los Seres HumanosLuis IguaranNo ratings yet
- TABAJO 1 MUSCULACION - Entrenamiento de La Fuerza. Editorial Médica Panamericana.Document8 pagesTABAJO 1 MUSCULACION - Entrenamiento de La Fuerza. Editorial Médica Panamericana.Kevin DvNo ratings yet
- El Tipo de Trabajo Muscular y Su Influencia en La FunciónDocument22 pagesEl Tipo de Trabajo Muscular y Su Influencia en La FunciónJuan Monsálvez SanhuezaNo ratings yet
- Adaptaciones fisiológicas del ejercicioDocument4 pagesAdaptaciones fisiológicas del ejercicioFelipe IzquierdoNo ratings yet
- La Función Muscular en La Abreacción TerapéuticaDocument49 pagesLa Función Muscular en La Abreacción TerapéuticaRenee CollinsNo ratings yet
- Estiramientos Mio-Tendino - Aponeuroticos 2 PDFDocument12 pagesEstiramientos Mio-Tendino - Aponeuroticos 2 PDFAnnieNo ratings yet
- Fundamentos de La Electromiografía de Superficie Aplicados A La Rehabilitación Física y BiomecánicaDocument3 pagesFundamentos de La Electromiografía de Superficie Aplicados A La Rehabilitación Física y BiomecánicaMaki GuillénNo ratings yet
- Sistema MuscularDocument19 pagesSistema MuscularÁlex LomelíNo ratings yet
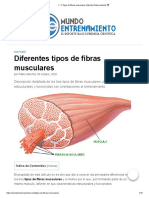
- Tipos de Fibras Musculares Características y FunciónDocument8 pagesTipos de Fibras Musculares Características y Funciónyelitza marinNo ratings yet
- La FlexibilidadDocument20 pagesLa FlexibilidadFederico HlfNo ratings yet
- FISIOLOGÍA DEL EJERCICIO Luis JavierDocument6 pagesFISIOLOGÍA DEL EJERCICIO Luis JavierDavid CameloNo ratings yet
- Lesiones EspDocument12 pagesLesiones EspSaid CuevasNo ratings yet
- Tema 2013 El Sistema MuscularDocument5 pagesTema 2013 El Sistema MuscularangelgadNo ratings yet
- Tema 4-3Document35 pagesTema 4-3MARIA DEL ROSARIO DUARTENo ratings yet
- Flexibilidad y componentes muscularesDocument13 pagesFlexibilidad y componentes muscularesManuel OvelarNo ratings yet
- .Document2 pages.nani longhiniNo ratings yet
- Mecánica muscular: contracción, tipos y propiedadesDocument31 pagesMecánica muscular: contracción, tipos y propiedadesDaniela GonzalesNo ratings yet
- Sistema MuscularDocument14 pagesSistema Muscularruz marina garrido De CastilloNo ratings yet
- Mecanica Del Sistema Oseo BIOFISICADocument7 pagesMecanica Del Sistema Oseo BIOFISICAQuimica Aplicada INo ratings yet
- Adaptaciones fisiológicas de la práctica del ejercicio físico: tipos de fibras muscularesDocument7 pagesAdaptaciones fisiológicas de la práctica del ejercicio físico: tipos de fibras muscularesFelipe IzquierdoNo ratings yet
- Fibras muscularesDocument8 pagesFibras muscularesabner romavi vargas uhNo ratings yet
- Tema 23 - Oposición EFDocument36 pagesTema 23 - Oposición EFVíctor Sánchez AguilarNo ratings yet
- McArdle NeuromuscularDocument95 pagesMcArdle NeuromuscularTomi CatalaNo ratings yet
- Ciencias ejercicio anatomía fisiologíaDocument2 pagesCiencias ejercicio anatomía fisiologíaCoachjd JiménezNo ratings yet
- Nsca - Manual de Entrenador.Document2 pagesNsca - Manual de Entrenador.Coachjd JiménezNo ratings yet
- SEPARATA Semana. #10.Document23 pagesSEPARATA Semana. #10.Jeremy Torres CartagenaNo ratings yet
- Biomecanica Del Tendon Revista Reduca PDFDocument13 pagesBiomecanica Del Tendon Revista Reduca PDFSebastián Vilches0% (1)
- MiologiaDocument6 pagesMiologiaDaniela SalinasNo ratings yet
- Cap 10Document6 pagesCap 10Raffi IstephanianNo ratings yet
- Trabajo Practico Musculacion - Fuerza - Ipef 2013Document6 pagesTrabajo Practico Musculacion - Fuerza - Ipef 2013Kevin DvNo ratings yet
- FUERZA 2021 Parte 1ADocument35 pagesFUERZA 2021 Parte 1ApaulaNo ratings yet
- Reparacion LigamentosDocument9 pagesReparacion LigamentosAlejandraRiveraLondoñoNo ratings yet
- Sistema Muscular - Wikipedia, La Enciclopedia LibreDocument12 pagesSistema Muscular - Wikipedia, La Enciclopedia LibremilagrosNo ratings yet
- Taller 5 Sistema Muscular 1810342 1810353Document6 pagesTaller 5 Sistema Muscular 1810342 1810353Bryan MarquezNo ratings yet
- BiomecánicaDocument14 pagesBiomecánicaLoNo ratings yet
- Los Músculos y La Unidad MotoraDocument14 pagesLos Músculos y La Unidad MotoraDoriany MtzNo ratings yet
- Tipos fibras muscularesDocument15 pagesTipos fibras muscularesRigoberto Jimenez GonzalezNo ratings yet
- Ergonomia AntropometriaDocument35 pagesErgonomia AntropometriaOrtiz CelinaNo ratings yet
- Contraccion Muscular Parte 2Document14 pagesContraccion Muscular Parte 2Gustavo GaleanoNo ratings yet
- 3.1 Estructura MuscularDocument12 pages3.1 Estructura MuscularSayra Lizzett Vargas VillalobosNo ratings yet
- Fisiologia de La Contraccion Muscular en El EjercicioDocument15 pagesFisiologia de La Contraccion Muscular en El EjercicioMaritza Romero ChuraNo ratings yet
- Biomecanica Del Musculo 2021Document21 pagesBiomecanica Del Musculo 2021Marina GámezNo ratings yet
- Fisiología del ejercicio físico y del entrenamientoFrom EverandFisiología del ejercicio físico y del entrenamientoRating: 4.5 out of 5 stars4.5/5 (8)
- ArtrosisDocument35 pagesArtrosisLyon DamiànNo ratings yet
- Presentacion EficazDocument2 pagesPresentacion EficazPriscila Suazo CornejoNo ratings yet
- A.T.M. Una Aproximacion Integrada de Tratamiento....Document5 pagesA.T.M. Una Aproximacion Integrada de Tratamiento....donjuandecasaNo ratings yet
- Marco TeóricoDocument13 pagesMarco TeóricoPriscila Suazo CornejoNo ratings yet
- A.T.M. Una Aproximacion Integrada de Tratamiento....Document5 pagesA.T.M. Una Aproximacion Integrada de Tratamiento....donjuandecasaNo ratings yet
- INTERSTICIODocument5 pagesINTERSTICIOGriNo ratings yet
- Guía completa para el dolor miofascialDocument718 pagesGuía completa para el dolor miofascialLaly Palacios MariñosNo ratings yet
- Seram2012 S-0366Document42 pagesSeram2012 S-0366ughoNo ratings yet
- Caso Clinico Consulta Ext.Document52 pagesCaso Clinico Consulta Ext.Andrea LunaNo ratings yet
- Tabulador Flex 2023 VFDocument216 pagesTabulador Flex 2023 VFErik BalderasNo ratings yet
- Abordaje Anterior de CaderaDocument20 pagesAbordaje Anterior de CaderaOciel AcostaNo ratings yet
- Musculacion y Sus EstiramientosDocument28 pagesMusculacion y Sus EstiramientosYoali TorresNo ratings yet
- Terapia ManualDocument33 pagesTerapia ManualCristina Huerta100% (3)
- ENERODocument84 pagesENEROjesus_salazar_ayestaranNo ratings yet
- Cupping TeraphyDocument4 pagesCupping TeraphyNancy GLEZNo ratings yet
- Anat PP Petros JOPDocument48 pagesAnat PP Petros JOPjorgeorihuela2478No ratings yet
- Necrectomia QuirurgicaDocument11 pagesNecrectomia QuirurgicaGiordan Sanchez100% (1)
- Diaterma HeidyDocument2 pagesDiaterma HeidyMilena Calderon guerreroNo ratings yet
- AbdomenDocument13 pagesAbdomenEric VillalobosNo ratings yet
- Compartimentos Musculares.Document5 pagesCompartimentos Musculares.heavenhell75No ratings yet
- Método RolfingDocument14 pagesMétodo RolfingCarlos Adrian Quintanilla VelascoNo ratings yet
- Desgarro MuscularDocument7 pagesDesgarro MuscularMariie McguireNo ratings yet
- Agentes físicos para fascitis plantarDocument3 pagesAgentes físicos para fascitis plantarBárbara RiquelmeNo ratings yet
- Abordaje Fisioterapico Del Sindrome Del Tunel Tarsiano Caso ClinicoDocument1 pageAbordaje Fisioterapico Del Sindrome Del Tunel Tarsiano Caso ClinicolyloteamoNo ratings yet
- Informe Practica ClaudiaDocument22 pagesInforme Practica ClaudiaAdam BoydNo ratings yet
- Fascia 1.en - EsDocument13 pagesFascia 1.en - EsPe T. ErNo ratings yet
- FasciasMúsculosCuelloDocument30 pagesFasciasMúsculosCuelloHector GuzmanNo ratings yet
- Fascitis PlantarDocument7 pagesFascitis PlantarEduardo Carvajal GonzálezNo ratings yet
- Liberación MiofascialDocument22 pagesLiberación MiofascialAndy Saritama100% (3)
- DupuytrenDocument6 pagesDupuytrenRaul Dennis Linares LinaresNo ratings yet
- Manual de AmputadosDocument41 pagesManual de AmputadosJavier Alonso Jara Cánovas100% (4)
- Terminología y Clasificación de Las Lesiones Musculares en El Deporte Concejo de Munich Articulo de Keily para Mapa ConceptualDocument31 pagesTerminología y Clasificación de Las Lesiones Musculares en El Deporte Concejo de Munich Articulo de Keily para Mapa ConceptualLizeth Dita PlataNo ratings yet
- Exposición Kinesio-TapeDocument17 pagesExposición Kinesio-Tapesebastian cardenasNo ratings yet
- Masaje Miofacial PDFDocument9 pagesMasaje Miofacial PDFJose Miguel Pompa Gil100% (2)
- Bordoni y Myers Feb 2020Document12 pagesBordoni y Myers Feb 2020beatrizhasanNo ratings yet