You might also like
- Herramientas As en Equilibrio Acido Base CorregidoDocument37 pagesHerramientas As en Equilibrio Acido Base CorregidoSarita DavilaNo ratings yet
- Balon de Contrapulsacion AorticaDocument32 pagesBalon de Contrapulsacion AorticaAngel Augusto Perez CalatayudNo ratings yet
- Anatomía cardíaca radiológicaDocument49 pagesAnatomía cardíaca radiológicaSilvana María Espinoza CuadrosNo ratings yet
- Rutina Masa MuscularDocument1 pageRutina Masa MuscularRodrigo Rodriguez ArnezNo ratings yet
- Stephan KorteDocument3 pagesStephan KorteGustavoNo ratings yet
- DeliriumDocument68 pagesDeliriumSarita DavilaNo ratings yet
- Formato AntropometriaDocument1 pageFormato AntropometriaHéctor OrtizNo ratings yet
- Ultrasonido pulmonarDocument16 pagesUltrasonido pulmonarDani Franco VargasNo ratings yet
- Fisiologia Cardiaca PostgradoDocument49 pagesFisiologia Cardiaca PostgradoDiego ManzaneroNo ratings yet
- 1proyecciones Radiológicas Del Corazón (Serie Cardiaca)Document35 pages1proyecciones Radiológicas Del Corazón (Serie Cardiaca)Oscar Segun DoNo ratings yet
- Historia ClinicaDocument36 pagesHistoria ClinicaAngela TovarNo ratings yet
- Cómo entrenar y alimentarse si tienes un cuerpo ectomorfoDocument3 pagesCómo entrenar y alimentarse si tienes un cuerpo ectomorfoANGELA SOTOMAYORNo ratings yet
- Órgano de ZuckerkandlDocument6 pagesÓrgano de ZuckerkandlAlexander MendezNo ratings yet
- Post-Operatorio de CCVDocument43 pagesPost-Operatorio de CCVPablo aguirre castilloNo ratings yet
- Glosario Radiológico Del TóraxDocument24 pagesGlosario Radiológico Del TóraxRolando ReynaNo ratings yet
- Segmentacion HepaticaDocument8 pagesSegmentacion HepaticaChrist FloreNo ratings yet
- Zonas de EntrenamientoDocument44 pagesZonas de EntrenamientoProfelaNo ratings yet
- TERMINOLOGIADocument7 pagesTERMINOLOGIAMaria Fernanda AngaritaNo ratings yet
- Presentacion DifusionDocument24 pagesPresentacion DifusionIvan Ignacio Subiabre TrujilloNo ratings yet
- Presentación TC de Cuello y ToraxDocument24 pagesPresentación TC de Cuello y ToraxAdalgisa GomezNo ratings yet
- Fisiología Cardiaca de Un DeportistaDocument14 pagesFisiología Cardiaca de Un DeportistaDaniela OrtizNo ratings yet
- Anatomia CorazonDocument14 pagesAnatomia CorazonMiguel M. Melchor RodríguezNo ratings yet
- Creatina, Uno de Los Mejores Suplementos. Beneficios, Usos, Dosis, Riesgos... Fitness RevolucionarioDocument1 pageCreatina, Uno de Los Mejores Suplementos. Beneficios, Usos, Dosis, Riesgos... Fitness RevolucionarioKlender100% (1)
- Criterios ECGDocument4 pagesCriterios ECGcentauro1119No ratings yet
- PletismografoDocument3 pagesPletismografovictor riosNo ratings yet
- Fisiologia Del EjercicioDocument9 pagesFisiologia Del EjercicioLorePazNo ratings yet
- La Calistenia y La Frecuencia CardiacaDocument4 pagesLa Calistenia y La Frecuencia Cardiacalaura garciaNo ratings yet
- Hemodinamia BasicaDocument4 pagesHemodinamia Basicajhon_paz7No ratings yet
- Frecuencia Cardiaca PulsoDocument8 pagesFrecuencia Cardiaca PulsoronalfmxNo ratings yet
- RADIOGRAFIA DE TORAX FinalDocument41 pagesRADIOGRAFIA DE TORAX FinalPatricia AceroNo ratings yet
- Algoritmos RCP AHA 2010Document16 pagesAlgoritmos RCP AHA 2010poroto17No ratings yet
- Bases Del ElectrocardiogramaDocument42 pagesBases Del ElectrocardiogramaEdith Hernandez FloresNo ratings yet
- Arteriografía femoral: estudio radiológico de la circulación arterial inferiorDocument12 pagesArteriografía femoral: estudio radiológico de la circulación arterial inferiorMelissa Ramirez100% (2)
- Calculo Correcto y Exacto de Balance Hidrico en AdultosDocument27 pagesCalculo Correcto y Exacto de Balance Hidrico en AdultosMauricio M Kramer100% (1)
- Trabajo EcgDocument14 pagesTrabajo EcgPau Torres SepúlvedaNo ratings yet
- ECOGRAFÍADocument3 pagesECOGRAFÍANacho InfanteNo ratings yet
- Rutina WeiderDocument7 pagesRutina WeiderKarlos EstévezNo ratings yet
- Caso clínico COVID cardíacoDocument20 pagesCaso clínico COVID cardíacoGiuliana MenachoNo ratings yet
- Tresguerres Capitulo 38 - FisiologiaDocument37 pagesTresguerres Capitulo 38 - FisiologiaEric KirNo ratings yet
- TP Fisiología N°4-Ley de Starling en El Corazón de SapoDocument7 pagesTP Fisiología N°4-Ley de Starling en El Corazón de SapoCecilia PascoNo ratings yet
- Cardiologia RVMDocument19 pagesCardiologia RVMapi-472592451No ratings yet
- Taller 3 FisiologíaDocument15 pagesTaller 3 FisiologíaANA LUISA ORTEGA LOPEZNo ratings yet
- ARRITMIASDocument21 pagesARRITMIASJoselyn RodriguezNo ratings yet
- Efecto de La Temperatura Sustancias en La Funcion Cardiaca en AnfibiosDocument8 pagesEfecto de La Temperatura Sustancias en La Funcion Cardiaca en AnfibiosAngelManriqueCristobalNo ratings yet
- Tarea sapITODocument3 pagesTarea sapITOGabriel DunasNo ratings yet
- Des Fisiologicas Del Musculo CardiacoDocument11 pagesDes Fisiologicas Del Musculo CardiacoANTHONYNo ratings yet
- Musculo Cardiaco WordDocument11 pagesMusculo Cardiaco WordRoberto Antonio Fiestas AmayaNo ratings yet
- Músculo cardíaco del sapo BufoDocument14 pagesMúsculo cardíaco del sapo BufoBriza Paulina CordovaNo ratings yet
- Actividad Eléctrica Del CorazónDocument44 pagesActividad Eléctrica Del CorazónMelany PaspuezanNo ratings yet
- CARDIORESPIDocument129 pagesCARDIORESPIAndrea OreaNo ratings yet
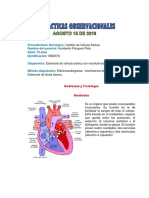
- Práctica Fisiología. Propiedades Del Corazón.Document13 pagesPráctica Fisiología. Propiedades Del Corazón.Berly Ruiz OrbezoNo ratings yet
- Contraccion Sistema ConduccionDocument63 pagesContraccion Sistema ConduccionUSMP FN ARCHIVOS100% (1)
- Fisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDocument6 pagesFisiología Del Musculo Cardiaco en Bufo Bufo Como Modelo ExperimentalDiego Jose Campos LeonNo ratings yet
- Reemplazo Valvula MitralDocument26 pagesReemplazo Valvula Mitralapi-394187731No ratings yet
- Comunicación InterauricularDocument12 pagesComunicación InterauricularDaniela MonsalveNo ratings yet
- YAFisiología Circulatoria II OSCAR PASTRANADocument37 pagesYAFisiología Circulatoria II OSCAR PASTRANACami ValerosNo ratings yet
- Práctica Disección Del CorazónDocument4 pagesPráctica Disección Del CorazónJuan TenjoNo ratings yet
- Clase 1 NefronaDocument5 pagesClase 1 NefronaGabriela AlvarezNo ratings yet
- Informe de Revascularizacion de MiocardioDocument12 pagesInforme de Revascularizacion de Miocardioapi-498610718No ratings yet
- Cambio de Valvula AorticaDocument8 pagesCambio de Valvula AorticaViviana SepulvedaNo ratings yet
- Morfologia Interna de Un InsectoDocument41 pagesMorfologia Interna de Un InsectoDEICY VANESSA RODRIGUEZ BARONNo ratings yet
- FinalDocument383 pagesFinallittleangel13471632No ratings yet
- Control de la presión arterialDocument21 pagesControl de la presión arterialHalabi Mehetani Ijab HamadNo ratings yet
- CARDIOPATIADocument25 pagesCARDIOPATIADanielaNo ratings yet
- Act2 LopezGuardadoDocument10 pagesAct2 LopezGuardadoLuis Antonio Lopez GuardadoNo ratings yet
- Anomalias CardiacasDocument6 pagesAnomalias CardiacasGermanNo ratings yet
- Taller 2 Riesgo ElectricoDocument4 pagesTaller 2 Riesgo ElectricoAndres GuzmanNo ratings yet
- Curso técnico de desfibriladoresDocument16 pagesCurso técnico de desfibriladorescarlosNo ratings yet
- Anatomia y Fisiologia IiDocument208 pagesAnatomia y Fisiologia Iimilagros palacios caroNo ratings yet
- Guia de - Sistema de ConduccionDocument6 pagesGuia de - Sistema de Conduccionapi-522436743No ratings yet
- TP4 EsplacnoDocument27 pagesTP4 EsplacnopabloNo ratings yet
- Folleto MasajeDocument67 pagesFolleto MasajeRodobaldo Mojena0% (1)
- Protocolo de Disección de Corazón de CerdoDocument10 pagesProtocolo de Disección de Corazón de CerdoSanjuan Cruz ValeriaNo ratings yet
- Berrospi JosephDocument4 pagesBerrospi JosephJoseph Alexander Berrospi FigueroaNo ratings yet
- Excitacion Ritmica Del CorazonDocument20 pagesExcitacion Ritmica Del CorazonAnonymous KoYsIn100% (1)
- CardioDocument17 pagesCardioJuan Ca DuarteNo ratings yet
- Delfín RosadoDocument4 pagesDelfín RosadoJohanna GámezNo ratings yet
- Manual de Interpretacion de HolterDocument58 pagesManual de Interpretacion de HolterOtero100% (2)
- Taponamiento CardíacoDocument24 pagesTaponamiento CardíacoJesana RicañoNo ratings yet
- Tetralogía de FallotDocument13 pagesTetralogía de FallotGUSTAVO FRANCISCO CONCHA VALLADARESNo ratings yet
- Poster Algoritmo PCR Traumatica Espanol 2015Document1 pagePoster Algoritmo PCR Traumatica Espanol 2015EduLópezNo ratings yet
- 8°CIENCIAS4Document6 pages8°CIENCIAS4PAULA ANDREA VIDAL CÁRDENANo ratings yet
- Carro Rojo y CardioversionDocument46 pagesCarro Rojo y Cardioversionsara ruizNo ratings yet
- Trabajo Colaborativo Tarea2 Grupo56Document48 pagesTrabajo Colaborativo Tarea2 Grupo56Laura MoralesNo ratings yet
- Paciente Con Insuficiencia Renal Crónica Moderada y Descompensación Cardíaca SobreañadidaDocument7 pagesPaciente Con Insuficiencia Renal Crónica Moderada y Descompensación Cardíaca SobreañadidaNicolás TovarNo ratings yet
- Sindrome de Noonan GUIA S N RASOPATIASDocument60 pagesSindrome de Noonan GUIA S N RASOPATIASnicole de la cruzNo ratings yet
- Anatomia HumanaDocument10 pagesAnatomia HumanaHugo JavierNo ratings yet
- Determina espacios intercostales ECGDocument9 pagesDetermina espacios intercostales ECGDarwin HurtadoNo ratings yet
- TEMA 7. ENFERMEDADES DEL MIOCARDIO, ENDOCARDIO y PERICARDIODocument25 pagesTEMA 7. ENFERMEDADES DEL MIOCARDIO, ENDOCARDIO y PERICARDIOAna Belén Quintanilla JuanNo ratings yet
- Abc y ConstituciónDocument9 pagesAbc y ConstituciónRafael MataNo ratings yet