You might also like
- Modelamiento y casos especiales de la cinética química heterogéneaFrom EverandModelamiento y casos especiales de la cinética química heterogéneaRating: 3 out of 5 stars3/5 (1)
- Teoría Del Gas Real: La Relación Exacta Para Gases RealesFrom EverandTeoría Del Gas Real: La Relación Exacta Para Gases RealesNo ratings yet
- (Trabajo) Termodinámica de SolucionesDocument9 pages(Trabajo) Termodinámica de SolucionesPaikuhan JoseNo ratings yet
- Wuolah-Capítulo 4Document8 pagesWuolah-Capítulo 4ksaweryjpcNo ratings yet
- Potencial QuimicoDocument15 pagesPotencial QuimicoRicardoNo ratings yet
- Termodinámica I: Relaciones termodinámicasDocument23 pagesTermodinámica I: Relaciones termodinámicasomar caballeroNo ratings yet
- Energía Libre de HelmholtzDocument13 pagesEnergía Libre de HelmholtzLuis Fernando Figuera Arias100% (1)
- Apunte Termodinámica de SolucionesDocument10 pagesApunte Termodinámica de Solucionesmatias ponsNo ratings yet
- Sistemas MulticomponentesDocument19 pagesSistemas MulticomponentesJoe BoykaNo ratings yet
- Preguntas de FinalDocument5 pagesPreguntas de FinalM Sol FiloniNo ratings yet
- Termodinámica de DisolucionesDocument4 pagesTermodinámica de DisolucionesAngelEmilioSalazarNo ratings yet
- Termodinámica aplicaciones Keq, H° y SDocument10 pagesTermodinámica aplicaciones Keq, H° y SJaviera Navarrete RiverosNo ratings yet
- UntitledDocument20 pagesUntitledSthefanie RangelNo ratings yet
- Ensayounidad2 FisicoquimicaDocument12 pagesEnsayounidad2 FisicoquimicaMARGARITA MOO CHANNo ratings yet
- Termodinámica, enzimas y cinéticaDocument7 pagesTermodinámica, enzimas y cinéticaJose Ignacio BarrientosNo ratings yet
- Tarea TermoDocument13 pagesTarea TermodanielYT.No ratings yet
- TermoquimicaDocument20 pagesTermoquimicaSayuriNo ratings yet
- Lectura 11 Fisico QuimicoDocument14 pagesLectura 11 Fisico QuimicoRuben AuronNo ratings yet
- 1.relaciones EstequiométricasDocument18 pages1.relaciones EstequiométricasCamila 22 SzafranNo ratings yet
- Conceptos Basicos en Los Procesos IndustrialesDocument27 pagesConceptos Basicos en Los Procesos IndustrialesPamela MartinezNo ratings yet
- UNI-México-FES-Aragón-IngEléctricaDocument17 pagesUNI-México-FES-Aragón-IngEléctricaEduardo PeñaNo ratings yet
- Sistemas Termicos 1Document3 pagesSistemas Termicos 1herichuNo ratings yet
- DocumentoDocument15 pagesDocumentobackyardigansjpcNo ratings yet
- Clase 3. Soluciones Ideales No IdealesDocument6 pagesClase 3. Soluciones Ideales No IdealesGloria Beatriz Delvalle MNo ratings yet
- Práctica 3 - TermodinamicaDocument7 pagesPráctica 3 - TermodinamicaLópez Jiménez CitlalliNo ratings yet
- Transformaciones y Cambios de Energía en Los Organismos y Sistemas Vivos - 070055Document9 pagesTransformaciones y Cambios de Energía en Los Organismos y Sistemas Vivos - 070055Juan David LatinezNo ratings yet
- Definicion y Algunos Ejercicios de Chang Segunda ParteDocument6 pagesDefinicion y Algunos Ejercicios de Chang Segunda ParteNohelia Barros Robles0% (1)
- Influencia Del Efecto Salino en La Velocidad de ReaccionesDocument36 pagesInfluencia Del Efecto Salino en La Velocidad de ReaccionesCris Hidalgo100% (2)
- Trabajo de EntalpiaDocument10 pagesTrabajo de EntalpiaHeidy Jovana Huanca RamosNo ratings yet
- La FugocidadDocument9 pagesLa FugocidadaddddddddfffffNo ratings yet
- Equilibrio Quimico de Sistemas HomogeneosDocument19 pagesEquilibrio Quimico de Sistemas HomogeneosKatia Yesica Tineo Canales0% (1)
- Potencial QuímicoDocument13 pagesPotencial QuímicoSandi Ham'GmzNo ratings yet
- Balance de Materia y EnergiaDocument12 pagesBalance de Materia y EnergiaGamaliel RamNo ratings yet
- Termodinamica de Las Reacciones QuimicasDocument30 pagesTermodinamica de Las Reacciones QuimicasDiana Paola BärränteNo ratings yet
- FQ BancopreguntasDocument8 pagesFQ BancopreguntasMax Salas100% (1)
- Resumen de Fisicoquimica 20Document13 pagesResumen de Fisicoquimica 20jennychoqueureyNo ratings yet
- Fugacidad y su relación con la ecuación de estadoDocument9 pagesFugacidad y su relación con la ecuación de estadoLuis MoreyNo ratings yet
- Bitacora 4 Frausto Sánchez KarlaDocument11 pagesBitacora 4 Frausto Sánchez Karlakarla fraustoNo ratings yet
- MEZCLASDocument5 pagesMEZCLASCarla Lara SalgadoNo ratings yet
- Termodinamica Trabajo VIDocument27 pagesTermodinamica Trabajo VIYexibel AguilarNo ratings yet
- Cap 4 Traducido CompletoDocument10 pagesCap 4 Traducido CompletoDiego QuinterosNo ratings yet
- Trabajo de Laboratorio P-7 Y P-8Document8 pagesTrabajo de Laboratorio P-7 Y P-8Guadalupe CallisayaNo ratings yet
- Influencia Del Efecto Salino en La Velocidad de ReaccionesDocument36 pagesInfluencia Del Efecto Salino en La Velocidad de Reaccioneslismarnieves1No ratings yet
- Fisico Soluciones RealesDocument26 pagesFisico Soluciones RealesAhilton SCNo ratings yet
- La Constante de EquilibrioDocument6 pagesLa Constante de EquilibrioCarlos RamírezNo ratings yet
- CaldeDocument8 pagesCaldeSamir GalanNo ratings yet
- Termodinámica y Diagramas de FaseDocument40 pagesTermodinámica y Diagramas de FaseJoaquin Patricio Valenzuela AstorgaNo ratings yet
- FisicoquimcaDocument6 pagesFisicoquimcasharon moreiraNo ratings yet
- Tarea 1 3 Er Parcial EntropiaDocument15 pagesTarea 1 3 Er Parcial EntropiaPaul Pineda QuinterosNo ratings yet
- Tema 2 BioqDocument13 pagesTema 2 BioqAlexandra María Marín BernalNo ratings yet
- Equilibrio fasesDocument7 pagesEquilibrio fasesSergioCastilloNo ratings yet
- Trabajo CorrosionDocument14 pagesTrabajo CorrosionGwynnbleidNo ratings yet
- Triangulacion 4Document21 pagesTriangulacion 4erickNo ratings yet
- Sistemas Multicomponentes Grupo 02 Trabajo FinalDocument26 pagesSistemas Multicomponentes Grupo 02 Trabajo FinalAllison JuarezNo ratings yet
- Termo Challa Pero Muy ChallaDocument4 pagesTermo Challa Pero Muy ChallaAgustín Soto DíazNo ratings yet
- Energía Libre de GibbsDocument10 pagesEnergía Libre de GibbsAlberto López100% (1)
- Estequiometría 4ESODocument4 pagesEstequiometría 4ESOChristian TorresNo ratings yet
- Formaulario PDFDocument2 pagesFormaulario PDFsyoran04No ratings yet
- Comenzar Con DropboxDocument4 pagesComenzar Con Dropboxvalsar33No ratings yet
- Metodo de ImagenesDocument7 pagesMetodo de ImagenesDavid A. LópezNo ratings yet
- Cla6 1Document6 pagesCla6 1syoran04No ratings yet
- ParametrizacionesDocument26 pagesParametrizacionesapi-3771554100% (4)
- Gradiente, Divergencia y RotacionalDocument7 pagesGradiente, Divergencia y RotacionalJose Francisco Hernandez OrtegaNo ratings yet
- TesisrfrangieDocument44 pagesTesisrfrangieBrenda Janeth Carrizosa RamosNo ratings yet
- Term Odin A MicaDocument1 pageTerm Odin A Micasyoran04No ratings yet
- Electromagnetism oDocument1 pageElectromagnetism osyoran04No ratings yet
- El Pendulo de Longitud Variable-LibreDocument4 pagesEl Pendulo de Longitud Variable-Libresyoran04No ratings yet
- Mecanica ClasicaDocument1 pageMecanica Clasicasyoran04No ratings yet
- Guia VimDocument116 pagesGuia Vimdrogs45No ratings yet
- CoordeadasDocument20 pagesCoordeadassyoran04No ratings yet
- hpf2 ApteDocument68 pageshpf2 Aptesyoran04No ratings yet
- Formato Carta de RecomendacionDocument3 pagesFormato Carta de Recomendacionsyoran04No ratings yet
- Creacion de Scripts en LinuxDocument32 pagesCreacion de Scripts en LinuxRoberto VenturaNo ratings yet
- George Gamow - El País de Las MaravillasDocument74 pagesGeorge Gamow - El País de Las Maravillassyoran04No ratings yet
- Einstein y Las MatematicasDocument8 pagesEinstein y Las MatematicasbetofisicosudNo ratings yet
- QuizDocument8 pagesQuizErika ZapataNo ratings yet
- Catalogo Productos RedVerdeDocument126 pagesCatalogo Productos RedVerdeAdolfo GoferNo ratings yet
- Algas CianofitasDocument13 pagesAlgas CianofitasFelipe PinzonNo ratings yet
- Interacciones Microorganismos Con Medio AmbienteDocument23 pagesInteracciones Microorganismos Con Medio AmbienteJosdany ChongNo ratings yet
- Examen de biología para estudiantes de secundariaDocument2 pagesExamen de biología para estudiantes de secundariaRos Bet ReyNo ratings yet
- Silabo BC 2018-IDocument23 pagesSilabo BC 2018-IPaulo Cesar Fernandez ArevaloNo ratings yet
- DavidAntónAlzamora1ºBAT CDocument2 pagesDavidAntónAlzamora1ºBAT CDavid A.ANo ratings yet
- Practica 5 de Biologia - CompressedDocument17 pagesPractica 5 de Biologia - CompressedMari SanchezNo ratings yet
- El par biomagnético: cómo tratar enfermedades con imanesDocument12 pagesEl par biomagnético: cómo tratar enfermedades con imanesRodrigo HMNo ratings yet
- Trabajo Pseudomona Aeroginosa.Document21 pagesTrabajo Pseudomona Aeroginosa.urielNo ratings yet
- Guía Membrana y Transporte 50calificación Etapa 6Document3 pagesGuía Membrana y Transporte 50calificación Etapa 6Denisse GalvezNo ratings yet
- Generalidades HongosDocument2 pagesGeneralidades Hongosfrancisco lozano gomezNo ratings yet
- Banco de Preguntas HistologiaDocument20 pagesBanco de Preguntas HistologiaMarco Martinez100% (1)
- 7 Ejemplos de Justficacion de Un ProyectoDocument12 pages7 Ejemplos de Justficacion de Un ProyectoJose KeyNo ratings yet
- PotencialDocument2 pagesPotencialErick HuertasNo ratings yet
- Guìa 5 Grado 6º Ciencias NaturalesDocument3 pagesGuìa 5 Grado 6º Ciencias NaturalesgenidNo ratings yet
- Derrames en Cavidades SerosasDocument10 pagesDerrames en Cavidades SerosasSonia Diaz GarciaNo ratings yet
- Biologia 8Document4 pagesBiologia 8Maria Fernanda Escudero BenenzianoNo ratings yet
- Examen Funcionamiento Celular Bases BiologicasDocument5 pagesExamen Funcionamiento Celular Bases Biologicasstefania LogronoNo ratings yet
- Anatomía de La FasciaDocument4 pagesAnatomía de La FasciaMirtaNo ratings yet
- Teoría atómica Dalton y origen elementos cuerpoDocument8 pagesTeoría atómica Dalton y origen elementos cuerpoShirley RodriguezNo ratings yet
- UT2 - Actividades RepasoDocument2 pagesUT2 - Actividades RepasoMiguel Ángel Campos RosaNo ratings yet
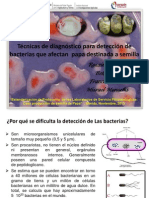
- Estandarización de Técnicas de Diagnóstico de Bacterias en Semilla de PapaDocument22 pagesEstandarización de Técnicas de Diagnóstico de Bacterias en Semilla de PapaJesus GarciaNo ratings yet
- Transporte de La MenbranaDocument6 pagesTransporte de La MenbranaMarycel ContrerasNo ratings yet
- Ciencias PruebaDocument5 pagesCiencias PruebaJuana BalagueraNo ratings yet
- Dieta y CáncerDocument45 pagesDieta y CáncerKEVIN GABRIEL ZU�IGA MOREIRANo ratings yet
- Guía Didactica N. 2.ciencias Nat. 7-1Document5 pagesGuía Didactica N. 2.ciencias Nat. 7-1Emy yayNo ratings yet
- Membrana Celular.Document51 pagesMembrana Celular.AXSSNo ratings yet
- Origen y evolución de los primeros reptilesDocument25 pagesOrigen y evolución de los primeros reptilesMoises Eliud DueñasNo ratings yet
- Celulas Madre y El CancerDocument28 pagesCelulas Madre y El CancerJeffry Noa MarínNo ratings yet