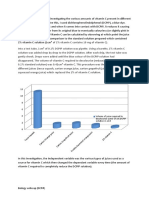
You might also like
- Invertase Session 3Document14 pagesInvertase Session 3Tiyah TimothyNo ratings yet
- LSM1102 - Practical 3 Lab Report (FULL)Document7 pagesLSM1102 - Practical 3 Lab Report (FULL)givena2ndchance100% (1)
- Determination of The Activity of InvertaseDocument7 pagesDetermination of The Activity of InvertaseVikki Anne CedoNo ratings yet
- Lab Report Water Potential FinalDocument6 pagesLab Report Water Potential Finalade2001fr3113No ratings yet
- Study Casein in Milk: Quantity of Casein ProteinDocument28 pagesStudy Casein in Milk: Quantity of Casein ProteindarshnaNo ratings yet
- Measure Leaf Transpiration Using a PotometerDocument3 pagesMeasure Leaf Transpiration Using a PotometerJeruel Saunders0% (1)
- Determination of The Solute Potential of Cell SapDocument4 pagesDetermination of The Solute Potential of Cell SapIshira Liyanage100% (1)
- The Vitamin C Content of Fruit JuiceDocument2 pagesThe Vitamin C Content of Fruit JuiceblasweedNo ratings yet
- Determination of Water Hardness by Complexometric Titration Class NotesDocument6 pagesDetermination of Water Hardness by Complexometric Titration Class NotesMehrnoosh Aref50% (2)
- Calculations TutorialDocument9 pagesCalculations Tutorialricara alexia moodley0% (1)
- The Extraction of Benzoic AcidDocument7 pagesThe Extraction of Benzoic AcidChenling NiNo ratings yet
- Biuret AssayDocument6 pagesBiuret Assay门门No ratings yet
- Determination of Ascorbic Acid Content of Some Fruit JuicesDocument17 pagesDetermination of Ascorbic Acid Content of Some Fruit JuicesReksy WibowoNo ratings yet
- Cyclohexanone Oxime Synthesis NotesDocument4 pagesCyclohexanone Oxime Synthesis NotesSherlock Wesley ConanNo ratings yet
- Bradford ReportDocument7 pagesBradford ReportNOXOLO NOMBULELO WENDY NGXONGONo ratings yet
- Comparing The Degree of Unsaturation of Olive Oil and That of Margarine Jameela Comments by FM PDFDocument2 pagesComparing The Degree of Unsaturation of Olive Oil and That of Margarine Jameela Comments by FM PDFFrank Massiah0% (1)
- Assay of Tissue GlycogenDocument26 pagesAssay of Tissue GlycogenTiyah TimothyNo ratings yet
- Enzymatic Browning and Its Prevention-American Chemical Society (1995)Document340 pagesEnzymatic Browning and Its Prevention-American Chemical Society (1995)danielguerinNo ratings yet
- Determination of Vitamin C in FoodsDocument11 pagesDetermination of Vitamin C in FoodsDalal Shab JakhodiyaNo ratings yet
- Determination of Vitamin C Lab ReportDocument3 pagesDetermination of Vitamin C Lab ReportMuhammadHasanAnsariNo ratings yet
- Chemistry 232 Determination of Vitamin C by An Iodometric TitrationDocument3 pagesChemistry 232 Determination of Vitamin C by An Iodometric TitrationHindami NugrohoNo ratings yet
- Carbs: Monosaccharides, Disaccharides & PolysaccharidesDocument6 pagesCarbs: Monosaccharides, Disaccharides & PolysaccharidesJada HartNo ratings yet
- Quantitative Determination of Proteins Using Bradford MethodDocument2 pagesQuantitative Determination of Proteins Using Bradford Methodann_michelle7No ratings yet
- Protein TechniquesDocument13 pagesProtein TechniquesRendel GuevarraNo ratings yet
- CATALASE: Structure, Function, and RegulationDocument9 pagesCATALASE: Structure, Function, and RegulationJed Dumadag Dano100% (3)
- Investigating Vitamin C Concentration in Homemade and Store Bought SmoothiesDocument7 pagesInvestigating Vitamin C Concentration in Homemade and Store Bought SmoothiesisabelleNo ratings yet
- Factors That Influence Enzyme Lab ActivityDocument5 pagesFactors That Influence Enzyme Lab ActivityDaniel BelnapNo ratings yet
- Vitamin C in FruitDocument10 pagesVitamin C in FruitWoon BingNo ratings yet
- Expt 3&4 - Peroxidase From RadishDocument9 pagesExpt 3&4 - Peroxidase From RadishLogeetha Geetha100% (1)
- TOPIC: Biological Molecules AIM: Identification of Biochemicals in Pure FormDocument46 pagesTOPIC: Biological Molecules AIM: Identification of Biochemicals in Pure FormManmohan SinghNo ratings yet
- Biochem 10A Lab QuestionsDocument6 pagesBiochem 10A Lab QuestionsPaul A IBattledaily Scavella100% (1)
- Spectrophotometric analysis of caffeine and benzoic acid in soft drinksDocument2 pagesSpectrophotometric analysis of caffeine and benzoic acid in soft drinksFeras SwaidNo ratings yet
- Simultaneous Determination of Salicylic Acid and Acetylsalicylic AciDocument5 pagesSimultaneous Determination of Salicylic Acid and Acetylsalicylic Aciiabureid7460No ratings yet
- Lab Manual Metal Acetylacetonate Complexes Web PDFDocument22 pagesLab Manual Metal Acetylacetonate Complexes Web PDFYan Jie ChongNo ratings yet
- Acidic and Basic Character of Organic CompoundsDocument35 pagesAcidic and Basic Character of Organic CompoundsLoveena Steadman100% (1)
- Measure Oxygen Uptake by RespirometerDocument17 pagesMeasure Oxygen Uptake by RespirometerTootsie100% (12)
- Column and Thin Layer ChromatographyDocument3 pagesColumn and Thin Layer ChromatographyChristine Evan HoNo ratings yet
- 1.vitamin C ContentDocument7 pages1.vitamin C ContentTheresa NgNo ratings yet
- Food TestsDocument7 pagesFood TestsDaneilla BanksNo ratings yet
- Measuring Potato Strip Water Potential Using Sucrose SolutionsDocument5 pagesMeasuring Potato Strip Water Potential Using Sucrose SolutionsSueEe100% (1)
- DCPIP Write UpDocument2 pagesDCPIP Write UpHashim Chishty0% (1)
- Estimation of Sugar in SoftdrinksDocument15 pagesEstimation of Sugar in SoftdrinksAhmadHabibBatubaraNo ratings yet
- Bio ReportDocument8 pagesBio ReportTharshini_Indr_6713No ratings yet
- Optimization of Extraction Conditions and Development of A SensitiveDocument6 pagesOptimization of Extraction Conditions and Development of A SensitiveSesilia Bernadet Soubirous MarshellaNo ratings yet
- Determination of Vitamin Content in Sample Tablets by TitrationDocument2 pagesDetermination of Vitamin Content in Sample Tablets by TitrationmagicianchemistNo ratings yet
- sbl1023 Lab 3 SpectrophotometerDocument6 pagessbl1023 Lab 3 Spectrophotometerapi-385146128No ratings yet
- Bio 100 Osmosis Lab ReportDocument4 pagesBio 100 Osmosis Lab Reportapi-249188694No ratings yet
- Browning ProcessDocument40 pagesBrowning ProcessRendy AndikaNo ratings yet
- AbstractDocument15 pagesAbstractAzzian AriffinNo ratings yet
- BCH 314 Tutorial 1 SolutionsDocument9 pagesBCH 314 Tutorial 1 SolutionsvictorNo ratings yet
- Class 11 Chemistry Notes 2023-24 8. Redox ReactionsDocument40 pagesClass 11 Chemistry Notes 2023-24 8. Redox ReactionsAyushi Shah100% (1)
- Enzyme KineticsDocument17 pagesEnzyme KineticsIsuru JayalathNo ratings yet
- UNIVERSITI TUNKU ABDUL RAHMAN Enzymology Experiment on Alkaline Phosphatase InhibitionDocument11 pagesUNIVERSITI TUNKU ABDUL RAHMAN Enzymology Experiment on Alkaline Phosphatase InhibitionCk Wong100% (1)
- 3.3 Determining K and V of Lactate Dehydrogenase: Table 3: Volumes Used For Dilution of 200mM Lactate (ML) For Tubes 1-5Document4 pages3.3 Determining K and V of Lactate Dehydrogenase: Table 3: Volumes Used For Dilution of 200mM Lactate (ML) For Tubes 1-5givena2ndchanceNo ratings yet
- Nutri Meta Lab 2 CombinedDocument16 pagesNutri Meta Lab 2 CombinedvlcjNo ratings yet
- Practical 3Document7 pagesPractical 3Gimme Your WafflesNo ratings yet
- Nutri Meta Lab 1Document11 pagesNutri Meta Lab 1vlcjNo ratings yet
- ENZYMOLOGYDocument41 pagesENZYMOLOGYbarbie.monster89No ratings yet
- Experiment 13 Ultrafiltration UnitDocument13 pagesExperiment 13 Ultrafiltration UnitKishen NaniNo ratings yet
- Experimental approaches to Biopharmaceutics and PharmacokineticsFrom EverandExperimental approaches to Biopharmaceutics and PharmacokineticsNo ratings yet
- Identification and Quantification of Secondary Metabolites .. - PDFDocument16 pagesIdentification and Quantification of Secondary Metabolites .. - PDFAmandaNo ratings yet
- Saudi Aramco Test Report: Internal Tank Bottom Testing 30-Oct-17 CP-SATR-X-3205Document8 pagesSaudi Aramco Test Report: Internal Tank Bottom Testing 30-Oct-17 CP-SATR-X-3205Tayyab AchakzaiNo ratings yet
- Activated CarbonDocument34 pagesActivated CarbonSanjeeva YedavalliNo ratings yet
- TestDocument94 pagesTestmutiaraNo ratings yet
- Progress in Polyethylene Terephthalate RecyclingDocument24 pagesProgress in Polyethylene Terephthalate RecyclingyuppeNo ratings yet
- Determination of Flash and Fire Point Using Cleveland ApparatusDocument10 pagesDetermination of Flash and Fire Point Using Cleveland ApparatusJawaria AhmedNo ratings yet
- ASTM D 5861-Ensayo de Adhesión FRP A Sustrato MetálicoDocument2 pagesASTM D 5861-Ensayo de Adhesión FRP A Sustrato MetálicoFernando Alfredo Sepúlveda JiménezNo ratings yet
- Spray Application of Belzona 2009Document4 pagesSpray Application of Belzona 2009Mounir BoutaghaneNo ratings yet
- 1984 WhiteheadDocument19 pages1984 WhiteheadAnita yuliyantiNo ratings yet
- Heat TreatmentDocument53 pagesHeat TreatmentIqbal Muhammad HusainiNo ratings yet
- Indirect RestorationsDocument32 pagesIndirect RestorationsNaji Z. Arandi100% (1)
- 2009 - Turner, J. W. - Death of A Child From Topical DiphenhydramineDocument2 pages2009 - Turner, J. W. - Death of A Child From Topical DiphenhydramineMaria KandelaNo ratings yet
- Material Safety Data Sheet Konkreton XSDocument9 pagesMaterial Safety Data Sheet Konkreton XSDaniel DaiaNo ratings yet
- Cyanide-Free Gold Ore Dressing Agent PDFDocument13 pagesCyanide-Free Gold Ore Dressing Agent PDFAldemar Arciniegas100% (1)
- Spent Coffee Grounds Heat Coffee RoastingDocument15 pagesSpent Coffee Grounds Heat Coffee RoastingMJundiNo ratings yet
- Distillation Tower DesignDocument66 pagesDistillation Tower DesignHavocFireNo ratings yet
- Dungs ATEX en 161 Solenoid ValvesDocument12 pagesDungs ATEX en 161 Solenoid Valvesstefan-romocki-4927No ratings yet
- 2012 A Level Answers P1 and P2 Compiled FinalDocument12 pages2012 A Level Answers P1 and P2 Compiled FinalWesley TanNo ratings yet
- FT8 MOBILEPAC Gas Turbine PackageDocument2 pagesFT8 MOBILEPAC Gas Turbine Packageudelmark100% (1)
- Astm A216-A216m-04Document4 pagesAstm A216-A216m-04NadhiraNo ratings yet
- How to Set Up a New Aquarium in 7 Easy StepsDocument40 pagesHow to Set Up a New Aquarium in 7 Easy StepsAndrew HardcastleNo ratings yet
- 1 MergedDocument8 pages1 MergedNo NameNo ratings yet
- THE THE: Notes Meter Factor ForDocument19 pagesTHE THE: Notes Meter Factor Forjgarcia388No ratings yet
- SATIP-A-114-01 Earthworks: Site Preparation, Excavation and Backfilling During ConstructionDocument2 pagesSATIP-A-114-01 Earthworks: Site Preparation, Excavation and Backfilling During ConstructionNino Celso AstilleroNo ratings yet
- GB T700-2006Carbonstructuralsteels (英文版)Document11 pagesGB T700-2006Carbonstructuralsteels (英文版)yong liNo ratings yet
- Maintain Your BRH 501 Demolition HammerDocument46 pagesMaintain Your BRH 501 Demolition HammerRomán Arturo Guerrero Uc100% (3)
- Air Pollution: by Bibhabasu MohantyDocument54 pagesAir Pollution: by Bibhabasu MohantyJaned PerwiraNo ratings yet
- Preparing Meds From Ampules and VialsDocument7 pagesPreparing Meds From Ampules and VialsZyra ObedencioNo ratings yet
- 0s SangDocument77 pages0s SangBijin PulikkottilNo ratings yet
- Hydrogen Fuel CellDocument27 pagesHydrogen Fuel CellVikash Mohanty100% (12)